
Россия, Калининград


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Была в сети 06.06.2018 00:05

Мудрицкая Светлана Викторовна
заместитель директора, учитель биологии
76 лет
Местоположение
Специализация
Учебное пособие "Мир клетки" Модуль 5 " Основные принципы строения эукариотической клетки" (продолжение) к Программе "Путешествие в мир клетки"
Категория:
Биология
28.05.2018 21:40
Учебник:
Биология. Общая биология. Профильный уровень. 10 класс. Захаров В.Б. и др. М.: 2010. - 352 с.

Просмотр содержимого документа
«Учебное пособие "Мир клетки" Модуль 5 " Основные принципы строения эукариотической клетки" (продолжение) к Программе "Путешествие в мир клетки"»
2.2.4.1. Ядерная оболочка
Структура, ограничивающая клеточное ядро - ядерная оболочка, характерна для эукариотических клеток. Она разделяет два внутриклеточных компартмента друг от друга, цитоплазму от ядра. Разделение этих структур в пространстве приводит к обособлению процессов синтеза белка и процессов синтеза нуклеиновых кислот, что создает дополнительные, по сравнению с прокариотами, возможности для регуляции генной активности. Активная регуляция транспорта из цитоплазмы в ядро и из ядра в цитоплазму через специальные комплексы пор создает систему избирательного транспорта веществ, делая ядерную оболочку «генными воротами», регулирующими потоки ядерного импорта и экспорта. Кроме того, ядерная оболочка играет большую роль в организации трехмерной структуры интерфазного ядра, элементы ядерной оболочки являются частью ядерного белкового матрикса.
Ядерная оболочка состоит из двух мембран, внешней и внутренней, между которыми располагается перинуклеарное пространство. Внутренняя мембрана ядерной оболочки структурно связана с ламиной - фиброзным периферическим слоем ядерного белкового матрикса. Ядерная оболочка имеет особые ядерные поры, которые образуются за счет многочисленных зон слияния двух ядерных мембран и представляют собой как бы округлые, сквозные перфорации всей ядерной оболочки.
В нешняя мембрана ядерной оболочки, непосредственно контактирующая с цитоплазмой клетки, связана с мембранной системой эндоплазматической сети (ЭПС), на ней обычно располагается большое количество рибосом, как и на мембранах эргастоплазмы. Существуют многочисленные наблюдения о непосредственном переходе внешней ядерной мембраны в систему каналов эндоплазматической сети.
нешняя мембрана ядерной оболочки, непосредственно контактирующая с цитоплазмой клетки, связана с мембранной системой эндоплазматической сети (ЭПС), на ней обычно располагается большое количество рибосом, как и на мембранах эргастоплазмы. Существуют многочисленные наблюдения о непосредственном переходе внешней ядерной мембраны в систему каналов эндоплазматической сети.
Внутренняя мембрана ядерной оболочки рибосом на своей поверхности не имеет, но связана с фиброзным слоем, ядерной ламиной, которая, в свою очередь, заякоревает хроматин на ядерной оболочке.
Фиброзный слой ламины все время перестраивается, особенно в связи с ростом поверхности ядра во время клеточного цикла. Характерные для внутренней ядерной мембраны белки ламины A, C и B относятся к фибриллярным белкам.
Наиболее характерной и бросающейся в глаза структурой в составе ядерной оболочки является ядерная пора. Поры в оболочке образуются за счет двух ядерных мембран в виде округлых сквозных отверстий или перфораций с диаметром около 100 нм. Округлое сквозное отверстие в ядерной оболочке заполнено сложно организованными глобулярными и фибриллярными структурами. Совокупность мембранных перфораций и этих структур называют комплексом пор ядра.
Я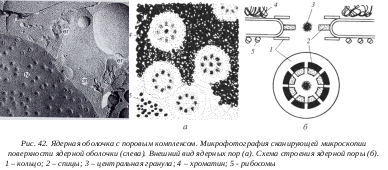 дерный поровый комплекс состоит из более 1000 белков, которые собраны примерно в 12 субкомплексов. Поровый комплекс представляет собой цилиндрическую фигуру. От периферических компонентов порового комплекса в сторону цитоплазмы простираются фибриллы белков. Со стороны ядра тоже фибриллярные выросты образуют корзинкоподобную структуру. Центр цилиндрической фигуры содержит «пробку» (центральную гранулу). Комплекс ядерной поры играет роль в контроле за ядерно-цитоплазменными связями. В процессе ядерно-цитоплазматического транспорта ядерные поры функционируют как некоторое молекулярное сито, пропуская частицы определенного размера пассивно, по градиенту концентрации. Так, ионы, сахара, нуклеотиды, АТФ, гормоны свободно поступают в ядра. С другой стороны ядерные поры осуществляют избирательный транспорт. Так, например, концентрация белков-гистонов в ядре значительно выше, чем в цитоплазме. Но, несмотря на это, во время синтеза ДНК происходит транспорт огромного количества (106 молекул каждые три минуты, или по 100-500 молекул через одну пору за 1 минуту) гистонов из цитоплазмы в ядро.
дерный поровый комплекс состоит из более 1000 белков, которые собраны примерно в 12 субкомплексов. Поровый комплекс представляет собой цилиндрическую фигуру. От периферических компонентов порового комплекса в сторону цитоплазмы простираются фибриллы белков. Со стороны ядра тоже фибриллярные выросты образуют корзинкоподобную структуру. Центр цилиндрической фигуры содержит «пробку» (центральную гранулу). Комплекс ядерной поры играет роль в контроле за ядерно-цитоплазменными связями. В процессе ядерно-цитоплазматического транспорта ядерные поры функционируют как некоторое молекулярное сито, пропуская частицы определенного размера пассивно, по градиенту концентрации. Так, ионы, сахара, нуклеотиды, АТФ, гормоны свободно поступают в ядра. С другой стороны ядерные поры осуществляют избирательный транспорт. Так, например, концентрация белков-гистонов в ядре значительно выше, чем в цитоплазме. Но, несмотря на это, во время синтеза ДНК происходит транспорт огромного количества (106 молекул каждые три минуты, или по 100-500 молекул через одну пору за 1 минуту) гистонов из цитоплазмы в ядро.
Из ядра через поры выходят в цитоплазму субъединицы рибосом и другие рибонуклеопротеиды. Эти данные показывают, что комплексы ядерных пор не представляют собой просто механические сита, которые ограничивают транспорт молекул в зависимости от их размеров.
2.2.4.2. Хроматин
Главный компонент ядер – это хроматин, выполняющий генетическую функцию клетки. В хроматиновой ДНК заложена практически вся генетическая информация. После окраски внутри ядра выявляются зоны плотного вещества, которое хорошо воспринимает разные красители, особенно основные. Благодаря такой способности хорошо окрашиваться этот компонент ядра и получил название «хроматин». Способность хроматина воспринимать основные красители определяется тем, что в состав хроматина входит ДНК в комплексе с белками.
Хроматин интерфазных ядер представляет собой несущие ДНК тельца, которые теряют в это время свою компактную форму, разрыхляются, деконденсируются. Степень такой деконденсации хромосом может быть различной в ядрах разных клеток. Когда хромосома или ее участок полностью деконденсирован, тогда эти зоны называют диффузным хроматином. При неполном разрыхлении хромосом в интерфазном ядре видны участки конденсированного хроматина.
С
тепень деконденсации хроматина в интерфазе может отражать функциональную активность этой структуры: чем более диффузен хроматин интерфазного ядра, тем выше в нем синтетические процессы транскрипции и репликации. Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде телец - хромосом, которые выполняют функцию распределения и переноса генетического материала в дочерние клетки.
Е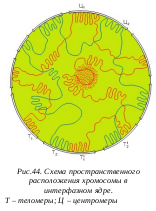 ще в начале 30-х годов XX века было замечено Э. Гейтцем, что в интерфазных ядрах существуют постоянные участки конденсированного хроматина, наличие которого не зависит от степени дифференцированнности ткани или от функциональной активности клеток. Такие участки получили название гетерохроматина, в отличие от остальной массы хроматина - эухроматина (собственно хроматина). Первоначально понятие гетерохроматина имело сугубо морфологическое значение, потому что при изучении препаратов окрашенных ядер нельзя знать, может ли данный участок конденсированного хроматина перейти в разрыхленное, эухроматическое состояние. На самом же деле только постоянно конденсированные участки хромосом в интерфазных ядрах сейчас принято называть гетерохроматином. Гетерохроматин генетически не активен, он не транскрибируется, реплицируется он позже всего остального хроматина. В состав гетерохроматина входит особая ДНК, обогащенная высокоповторяющимися последовательностями нуклеотидов; он локализован в центромерных, теломерных и интеркалярных зонах митотических хромосом. Функциональное значение гетерохроматина до конца не выяснено, предполагается, что он участвует в конъюгации хромосом во время мейоза.
ще в начале 30-х годов XX века было замечено Э. Гейтцем, что в интерфазных ядрах существуют постоянные участки конденсированного хроматина, наличие которого не зависит от степени дифференцированнности ткани или от функциональной активности клеток. Такие участки получили название гетерохроматина, в отличие от остальной массы хроматина - эухроматина (собственно хроматина). Первоначально понятие гетерохроматина имело сугубо морфологическое значение, потому что при изучении препаратов окрашенных ядер нельзя знать, может ли данный участок конденсированного хроматина перейти в разрыхленное, эухроматическое состояние. На самом же деле только постоянно конденсированные участки хромосом в интерфазных ядрах сейчас принято называть гетерохроматином. Гетерохроматин генетически не активен, он не транскрибируется, реплицируется он позже всего остального хроматина. В состав гетерохроматина входит особая ДНК, обогащенная высокоповторяющимися последовательностями нуклеотидов; он локализован в центромерных, теломерных и интеркалярных зонах митотических хромосом. Функциональное значение гетерохроматина до конца не выяснено, предполагается, что он участвует в конъюгации хромосом во время мейоза.
Основная масса хроматина ядра может менять степень своей компактизации в зависимости от функциональной активности, она относится к эухроматину. Эухроматические неактивные участки, которые находятся в конденсированном состоянии, стали называть факультативным гетерохроматином, подчеркивая необязательность такого его состояния. Хорошим примером факультативного гетерохроматина может служить X-хромосома в организме человека. В клетках мужской особи X-хромосома деконденсирована, она активна, транскрибируется и морфологически не выявляется из-за своего рыхлого, диффузного состояния. В клетках женского организма, где присутствуют две X-хромосомы, одна из них находится в активном, диффузном состоянии, а вторая - в неактивном, конденсированном, она временно гетерохроматизована. В этом состоянии она может существовать в течение всей жизни организма. Но потомки ее, попадая в клетки мужского организма следующего поколения, снова будут активированы.
В дифференцированных клетках всего лишь около 10% генов находится в активном состоянии, остальные гены инактивированы и, соответственно, находятся в составе конденсированного хроматина (факультативный гетерохроматин). Это обстоятельство объясняет, почему большая часть хроматина ядра структурирована.
2.2.4.3. Ядрышко
Внутри интерфазных ядер на окрашенных препаратах видны мелкие, обычно шаровидные тельца - ядрышки. Впервые ядрышки были описаны Фонтана в 1774 г. Ядрышки обнаруживаются практически во всех ядрах эукариотических клеток. В клеточном цикле ядрышко присутствует в течение всей интерфазы: в профазе оно постепенно исчезает, и отсутствует в мета- и анафазе и вновь появляется в середине телофазы.
В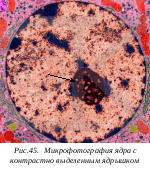 озникновение ядрышек связано с определенными зонами на особых, ядрышкообразующих участках хромосом. Эти зоны были названы ядрышковыми организаторами, а сами ядрышки представлялись как структурное выражение хромосомной активности.
озникновение ядрышек связано с определенными зонами на особых, ядрышкообразующих участках хромосом. Эти зоны были названы ядрышковыми организаторами, а сами ядрышки представлялись как структурное выражение хромосомной активности.
Уже в 50-х годах XX века при изучении ультраструктуры ядрышек в их составе были обнаружены гранулы, сходные по своим свойствам с цитоплазматическими гранулами рибонуклеопротеидной природы, с рибосомами. Следующим этапом в изучении ядрышка было открытие принципиального факта: «ядрышковый организатор» является вместилищем генов рибосомных РНК.
Внутренняя часть ядрышка имеет фибриллярную структуру, состоящую из участка ДНК – ядрышкового организатора и синтезирующихся нитей рРНК. Наружная гранулярная часть - это зона зрелых и созревающих субъединиц рибосом. Ядрышко участвует в образовании рибосом и мощно развито в клетках с интенсивным синтезом белка.
Ядерный матрикс состоит из трех морфологических компонентов: периферического белкового сетчатого слоя - лямины, внутреннего, или интерхроматинового матрикса, и «остаточного» ядрышка. Лямина образует сплошной фибриллярный слой, прилежащий к внутренней мембране кариолеммы, поддерживает морфологическую целостность ядра. Интерхроматиновый матрикс представлен рыхлой фибриллярной сетью, в ячейках которых располагается хроматин в разной стадии конденсации. К фибриллам присоединены комплексы ферментов синтеза нуклеиновых кислот. Остаточное ядрышко состоит из плотно уложенных фибрилл и повторяет форму ядрышка. Компоненты ядерного матрикса динамичны и меняются в зависимости от функциональных особенностей клеток.
При подготовке к практическому занятию по материалам учебного пособия продумайте, с участием каких органелл в клетке происходят следующие процессы:
| Процессы | Органеллы, клеточные структуры | Особенности ультраструктуры |
| Синтез липидов, холестерина | | |
| Синтез гликогена, полисахаридов | |
|
| Детоксикация ядов | |
|
| Транспорт веществ в клетке | |
|
| Синтез белков | |
|
| Формирование третичной и четвертичной структуры белка | | |
| Присоединение сахаров к белкам | |
|
| Образование лизосом | |
|
| Образование клеточных мембран | |
|
| Внутриклеточное пищеварение | |
|
| Процессы биологического окисления | |
|
| Хранение генетической информации | |
|
| Аккумуляция энергии солнца | |
|
| Образование рибосом | |
|
| Процессы транскрипции | |
|
| Процессы трансляции | |
|
| Обезврещивание Н2О2 | |
|
| Экзо – и эндоцитоз | |
|
| Формирование псевдоподий | |
|
| Передача нервного импульса с одного нейрона на другой | |
|
59













Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!
Полезное для учителя

Реализация образовательных программ осуществляется с применением исключительно электронного обучения и ДОТ

