
Россия, Республика Башкортостан, Уфа


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Была в сети 09.01.2024 14:50

Мустаева Лейсан Миннимухаматовна
учитель химии
Местоположение
Специализация
Литературный обзор: "Эволюция мозгового черепа древних рептилий. Какие из изображенных черепов принадлежат к анапсидному, синапсидному, диапсидному типам? Чем они отличаются друг от друга?"
Категория:
Биология
30.06.2017 12:14

Просмотр содержимого документа
«Литературный обзор: "Эволюция мозгового черепа древних рептилий. Какие из изображенных черепов принадлежат к анапсидному, синапсидному, диапсидному типам? Чем они отличаются друг от друга?"»
Эволюция мозгового черепа древних рептилий. Какие из изображенных черепов принадлежат к анапсидному, синапсидному, диапсидному типам? Чем они отличаются друг от друга?
Череп (cranium) – скелет головы позвоночных. Различают мозговой череп (cranium celebrale, s. neurocranium) и висцеральный (cranium viscerale, s. splanchnocranium). Череп пресмыкающихся видоизменялся главным образом в зависимости от характера питания и способов добывания пищи. Сплошной панцирь черепа подвергся частичной редукции. В связи с более дифференцированной мускулатурой челюстного аппарата в височной области черепа возникли ямы и разделяющие их костные мосты — дуги, служившие для прикрепления сложной системы мускулов. От черепа амфибий он отличается вытянутыми челюстями, образующими относительно длинное рыло. У пресмыкающихся дыхание обеспечивается работой грудной клетки, а захват добычи связан с активным преследованием, при котором вытянутое рыло имеет преимущество. Такая форма челюстей давала к тому же возможность отрывать куски от крупной добычи. То и другое требовало более мощной жевательной мускулатуры. С ее развитием и усложнением органов чувств связаны основные изменения черепа пресмыкающихся.
| Тип черепа | Строение | Характеристика | Характерные группы |
| Анапсидный |
| Череп без височных окон. | Парарептилии |
| Синапсидный |
| Пониженное положение височного окна, над которым смыкаются чешуйчатая и заднеглазничные кости. | Звероподобные рептилии |
| Диапсидный |
| Имеется два височных окна. | Лепидозавры и другие рептилии. |
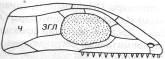
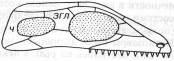
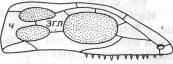
*ч – чешуйчатая кость, згл – заднеглазничная кость
1-ый тип - АНАПСИДНЫЙ. Самым главным достижением данного 1 типа явлется наличие формирования черепной ямки, к которой присоединяется жевательная мускулатура. (Анапсиды — с цельным черепным панцирем (котилозавры и черепахи).
2-ой тип - СИНАПСИДНЫЙ. Когда соответственно формируются верхняя и нижняя височная ямки.
К данному типу строения черепа можно отнести тереоморфов и тераморфов. (Синапсиды — с одной скуловой дугой (звероподобные, плезиозавры и, возможно, ихтиозавры).
3-ий тип - ДИАПСИДНЫЙ. Три височные ямки. (Диапсиды — с двумя дугами (все прочие пресмыкающиеся).
Какие позвоночные произошли от рептилий с диапсидным черепом,
какие дуги они при этом утратили?
Диапсидный череп имеет две височные впадины, расположенные выше и ниже заглазничной кости. Диапсидные в конце палеозоя (пермь) дали чрезвычайно широкую адаптивную радиацию систематическим группам и видам, которых обнаруживают и среди вымерших форм, и среди нынешних рептилий. Среди диапсидных наметились две основные группы Лепидозавроморфы (Lepidosauromorpha) и Архозавроморфы (Archosauromorpha). Наиболее примитивные диапсиды из группы Лепидозавров — отряд Эозухии (Eosuchia) — были предками отряда Клювоголовые, от которых в настоящее время сохранился лишь один род — гаттерия.
В конце перми от примитивных диапсид обособились чешуйчатые (Squamata), ставшие многочисленными в меловой период. К концу мелового периода от ящериц произошли змеи.
Какие позвоночные произошли от рептилий с синапсидным
черепом?
Cинапсиды — рептилии с нижними височными впадинами, ограниченными скуловой, чешуйчатой и заглазничной костями. Уже в позднем карбоне эта группа стала самой многочисленной. В палеонтологической летописи они представлены двумя последовательно существовавшими отрядами: пеликозавров (Pelicosauria) и терапсид (Therapsida). Период своего расцвета терапсиды пережили задолго до того, как появились первые динозавры, потомками терапсид стали более высокоорганизованные зверозубые (Theriodontia). К ним относится, в частности, хищная иностранцевия, напоминающая саблезубого тигра. Зверозубые были многочисленны ещё в раннем триасе, но после появления хищных динозавров они исчезли. Звероподобные представляют большой интерес как группа, давшая начало млекопитающим.
Чем вызвано образование в мозговом черепе височных ям?
В связи с воздействием разнообразных условий жизни, с которыми они сталкивались при размножении и расселении. Большинство групп приобрели бо́льшую подвижность; скелет их стал легче, но в то же время и прочнее. Рептилии использовали более разнообразную пищу, чем амфибии. Изменялась техника её добывания. В этой связи существенным изменениям подверглось строение конечностей, осевого скелета и черепа. У большинства конечности стали длиннее, таз, приобретая устойчивость, прикреплялся к двум и более крестцовым позвонкам. В плечевом поясе исчезла «рыбная» кость клейтрум. Сплошной панцирь черепа подвергся частичной редукции. В связи с более дифференцированной мускулатурой челюстного аппарата в височной области черепа возникли ямы и разделяющие их костные мосты — дуги, служившие для прикрепления сложной системы мускулов.
Литература:
1.Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989.
2.Баянов М.Г. 2003. Серая цапля в Башкортостане. – Итоги биологических исследований (Вып. 7): Сборник научных трудов. Уфа: 27-29.
3.Баянов М.Г., Валуев В.А. Большой баклан в Башкирии // Биоразнообразие и биоресурсы Урала и сопредельных территорий: Материалы II международной конференции. Оренбург: Изд-во ОГПУ, 2002. С. 150.
4.Валуев В.А. 2002. К авифауне лесостепи Предуралья Башкирии. – Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург: 69-71.
5.Валуев В.А. 2004. Экстраполяционный коэффициент как дополнение к учёту численности птиц по методике Ю.С. Равкина (1967) для территорий со значительной ландшафтной дифференциацией. – Вестник охотоведения, 1 (№3). М.: 291-293.
6.Валуев В.А. 2004. Обзор летней орнитофауны Предуралья и Южного Урала (отряды: гагарообразные, поганкообразные, веслоногие, аистообразные, фламингообразные, гусеобразные). – Вестник Башкирского университета. № 1. Уфа: 35-41.
7.Валуев В.А., Валуев К.В. 2001. Новые наблюдения редких птиц в Башкирии. – Итоги биологических исследований. Вып. 6. Сборник научных трудов. Уфа: 161-163.
8.Галиева Л.Ф. 2002. Интересные встречи птиц в Нагаево и на озере Кандры-куль в Башкирии. – Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург: 86-87.
9.Жизнь животных. Энциклопедия в шести томах. Том 5. (Земноводные. Пресмыкающиеся). Общая редакция члена-корреспондента АН СССР профессора Л. А. Зенкевича. — Москва: Просвещение, 1969. — 488 стр.
10.Жизнь животных: в 6-ти томах. — М.: Просвещение. Под редакцией профессоров Н.А.Гладкова, А.В.Михеева. 1970.
11.Животный мир Башкортостана/ под редакцией М. Г. Баянова и профессора Е. В. Кучерова/ Уфа, "КИТАП" 1995.
12.Книзе А. А. Применение маршрутного метода количественного учета боровой птицы в условиях Ленинградской области и АКССР. Сб. «Пром. фауна и охот, хоз.», Л. — М., 1934, стр. 27— 34.
13.Лилиенталь О. Полёт птиц как основа искусства летать. — М., 2002 (рус.)
14.Наумов Н. П., Карташев Н. Н. Часть 2. Пресмыкающиеся, птицы, млекопитающие // Зоология позвоночных. — М.: Высшая школа, 1979. — С. 272.
15.Новиков Г. А. Упрощенный метод количественного учета птиц. Природа, № 2, 1947, стр. 57.
16.НовиковГ.А."Полевые исследования экологии наземных позвоночных животных"
( изд. "Советская наука" 1949 г.)17.Семенов-Тян- Шанский О. И. Опыт учета боровой дичи в Лапландском заповеднике. Там же, стр. 55—59.
18.Стахровский В. Г. и Морин Н. А. Учет тетерева и рябчика. Сб. «Верхне-Вычегодск. эксп.», М., 1932, стр. 196—202.
19.Щербаков А. Ф. Тетерев-глухарь, его распространение и запас. Сб. «Верхне-Вычегодск. эксп.», М., 1932, стр. 148—162.
20.Якоби В. Э. Морфоэкологические приспособления к скоростному полёту у птиц. — Механизмы полёта и ориентации птиц. — М.: Наука, 1966.







Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!

