Россия, Анапа


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до 14.05.2025
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Был в сети 10.12.2022 15:58

Штрекер Иван Августович
Преподаватель
52 года
Местоположение
Специализация
История открытия ДНК
Категория:
Химия
15.11.2017 17:50
Тема:
§ 7. Полимеры

Просмотр содержимого документа
«История открытия ДНК»
Департамент образования и науки Кемеровской области
Муниципальное бюджетное общеобразовательное учреждение
«Средняя общеобразовательная школа №20»
РЕФЕРАТ
по химии
История открытия ДНК.
Выполнили:
учащиеся 11 класса:
Беккер Глеб
Власенко Алена
Гончар Софья
Темиртау
2017 год
Биография Дж. Уотсона
Джеймс Дьюи Уотсон родился 6 апреля 1928 года в Чикаго (штат Иллинойс). В родном городе мальчик получил начальное и среднее образование. Вскоре стало очевидно, что Джеймс необыкновенно одаренный ребенок, и его пригласили на радио для участия в программе «Викторины для детей». Лишь два года проучившись в средней школе, Уотсон получил в 1943 году стипендию для обучения в экспериментальном четырехгодичном колледже при Чикагском университете, где проявил интерес к изучению орнитологии. Окончив в 1947 году университет со степенью бакалавра естественных наук, он продолжил затем образование в Индианском университете Блумингтона.
К этому времени Уотсон заинтересовался генетикой и начал обучение в Индиане под руководством специалиста в этой области Г.Д. Меллера и бактериолога С. Лурия. В 1950 году молодой ученый получил степень доктора философии за диссертацию о влиянии рентгеновских лучей на размножение бактериофагов (вирусов, инфицирующих бактерии). Субсидия Национального исследовательского общества позволила ему продолжить исследования бактериофагов в Копенгагенском университете в Дании. Там он проводил изучение биохимических свойств ДНК бактериофага. Однако, как он позднее вспоминал, эксперименты с бактериофагом стали его тяготить, ему хотелось узнать больше об истинной структуре молекул ДНК, о которых так увлеченно говорили генетики.
В 1944 году Эйвери, Мак-Леод, Мак-Карти установили важнейший факт, что наследуемая передача признаков у бактерий осуществляется ДНК. Тогда же американский биолог О. Эйвери представил доказательства, что гены состоят из ДНК. Эту гипотезу подтвердили в 1952 году А. Херши и М. Чейз. Вместе с тем было ясно, что ДНК контролирует основные биохимические процессы, происходящие в клетке, но ни структура, ни функция молекулы не были известны.
Весной 1951 года, во время пребывания на симпозиуме в Неаполе, Уотсон встретил Мориса Уилкинса, английского исследователя. Уилкинс и Розалин Франклин, его коллега по Королевскому колледжу Кембриджского университета, провели рентгеноструктурный анализ молекул ДНК. В результате они доказали, что молекулы представляют собой двойную спираль, напоминающую винтовую лестницу. Полученные ими данные привели Уотсон к мысли исследовать химическую структуру нуклеиновых кислот. Национальное общество по изучению детского паралича выделило субсидию.
В октябре 1951 года ученый отправился в Кавендишскую лабораторию Кембриджского университета для исследования пространственной структуры белков совместно с Д.К. Кендрю. Там он и познакомился с Криком, физиком, интересовавшимся биологией и писавшим в то время докторскую диссертацию.
Несмотря на общность интересов, взглядов на жизнь и стиль мышления, Уотсон и Крик беспощадно, хотя и вежливо, критиковали друг друга. Их роли в этом интеллектуальном дуэте были разными. «Френсис был мозгом, а я - чувством», - говорит Уотсон.
Начиная с 1952 года, основываясь на ранних исследованиях Чаргаффа, Уилкинса и Франклин, Крик и Уотсон решили попытаться определить химическую структуру ДНК.
Вспоминая об отношении к ДНК подавляющего большинства биологов тех дней, Уотсон писал: «После опытов Эйвери было похоже, что именно ДНК основной генетический материал. Таким образом, выяснение химического строения ДНК могло оказаться важным шагом к пониманию того, как воспроизводятся гены. Но в отличие от белков, относительно ДНК имелось очень мало точно установленных химических сведений. Ею занимались считанные химики, и за исключением того факта, что нуклеиновые кислоты представляют собой очень большие молекулы, построенные из меньших строительных блоков - нуклеотидов, об их химии не было известно ничего такого, за что мог бы ухватиться генетик. Более того, химики-органики, работавшие с ДНК, почти никогда не интересовались генетикой».
Американские ученые постарались свести воедино все имевшиеся до сих пор сведения о ДНК, как физико-химические, так и биологические. Как пишет В.Н. Сойфер: «Уотсон и Крик подвергли анализу данные рентгеноструктурного анализа ДНК, сопоставили их с результатами химических исследований соотношения нуклеотидов в ДНК (правила Чаргаффа) и применили к ДНК идею Л. Полинга о возможности существования спиральных полимеров, высказанную им в отношении белков. В результате они смогли предложить гипотезу о структуре ДНК, согласно которой ДНК представлялась составленной из двух полинуклеотидных нитей, соединенных водородными связями и взаимно закрученных друг относительно друга. Гипотеза Уотсона и Крика так просто объясняла большинство загадок о функционировании ДНК как генетической матрицы, что она буквально сразу была принята генетиками и в короткий срок экспериментально доказана».
После опубликования описания модели в английском журнале «Нейче» в апреле 1953 года тандем Крика и Уотсона распался.
Через год с небольшим Уотсон был назначен старшим научным сотрудником кафедры биологии Калифорнийского технологического института в Пасадене (штат Калифорния). В 1955 году, когда он работал ассистентом профессора биологии в Гарвардском университете Кембриджа (штат Массачусетс), судьба вновь свела его с Криком, с которым он проводил совместные исследования до 1956 года. В 1958 году Уотсон был назначен адъюнкт-профессором, а в 1961 году - полным профессором.
Биография Ф. Крика
Старший из двух сыновей, Фрэнсис, родился в семье Гарри Крика и Элизабет Энн Уилкинс 8 июня 1916 года в Нортгемптоне, Англия. Он учился в местной гимназии и уже в раннем возрасте увлекся экспериментами, часто сопровождавшимися химическими взрывами. В школе он получил приз за сбор полевых цветов. Кроме того, он был помешан на теннисе, но не сильно интересовался другими играми и видами спорта. В возрасте 14 лет Фрэнсис получил стипендию школы Милл-Хилл на севере Лондона. Четыре года спустя, в 18 лет, он поступил в университетский колледж. К его совершеннолетию родители переехали из Нортгемптона в Милл-Хилл, и это позволило Фрэнсису во время учебы жить дома. Он получил диплом с отличием в области физики.
После бакалавриата Фрэнсис Крик под руководством да Коста Андраде в университетском колледже занимался исследованиями вязкости воды под давлением и при высоких температурах. В 1940 г. Фрэнсис получил гражданскую должность в Адмиралтействе, где работал над проектированием противокорабельных мин. В начале года Крик женился на Рут Дорин Додд. Их сын Майкл родился во время воздушного налета на Лондон 25 ноября 1940 года.
Во время Второй мировой войны он работал в научно-исследовательской лаборатории ВМС. Крик занимался разработкой магнитных и акустических мин и участвовал в проектировании новых типов мин, остающихся невидимыми для немецких кораблей-тральщиков (корабли специального назначения для обнаружения и удаления препятствий в виде мин).
К концу войны Фрэнсис был приписан к научной разведке в штаб-квартире британского Адмиралтейства в Уайтхолле, где занимался разработкой оружия.
Понимая, что ему потребуется дополнительное обучение, чтобы удовлетворить свое желание заняться фундаментальными исследованиями, Крик решил работать над ученой степенью. По его словам, он был очарован двумя областями биологии – границей между живым и неживым и деятельностью головного мозга. Крик выбрал первую, несмотря на то что мало знал о предмете. После предварительных исследований в университетском колледже в 1947 году он остановился на программе в лаборатории в Кембридже под руководством Артура Хьюза, касавшейся работы над физическими свойствами цитоплазмы культуры куриных фибробластов.
Два года спустя Крик присоединился к группе Совета по медицинским исследованиям в Кавендишской лаборатории. В нее вошли британские академики Макс Перуц и Джон Кендрю (будущие Нобелевские лауреаты). Фрэнсис стал с ними сотрудничать якобы для изучения структуры белка, но в действительности для работы с Уотсоном над разгадкой строения ДНК.
Предпосылки открытия ДНК
Проникая все глубже в тайны мироздания, человек пытался ответить на один из основных вопросов, которым задавались еще древние мудрецы: что есть жизнь, что есть сам человек? Тайна рождения живых организмов интересовала ученых не меньше, чем строение звезд. Открытия в области биологии, совершенные в XX в., вывели человечество на новые рубежи, наметили поистине фантастические перспективы. Молекулярная биология по‑прежнему остается одной из самых перспективных наук нашего времени.
Разработав теорию эволюции живых организмов, Дарвин не мог ответить на вопрос, как закрепляются в потомстве изменения в структуре и функциях живых организмов, возникшие в процессе этой эволюции. Но когда его книга только‑только вышла из печати, в Чехии уже ставил свои опыты Грегор Мендель . Его выводы положили начало развитию науки о наследственности – генетики, которой суждено было объяснить важнейшие загадки мироздания. На модели гороха Мендель впервые установил существование особых «наследственных факторов» (позднее названных «генами»), передающихся от одного поколения другому, переносящих при этом определенные признаки. Однако еще долгое время сам механизм передачи был ученым неизвестен.
В то же время в Германии работал зоолог Август Вейсман , который высказал и доказал правильность мнения о том, что переход родительских свойств на потомство зависит от прямой передачи родителями некоего материального вещества, которое, по мнению Вейсмана, было заключено в хромосомах – органеллах клетки. Важнейшие для развития генетики исследования в дальнейшем провел американец Томас Морган (см. приложения). Поставив массу экспериментов на мухах‑дрозофилах, он и его сотрудники пришли к выводам о материальных основах наследственности, линейной локализации генов в хромосомах, закономерности их мутационной изменчивости, цитогенетическом механизме их наследственной передачи и др., что позволило окончательно оформить основные принципы хромосомной теории наследственности.
В 1869 г. биохимик Мишер выделил из клеточных ядер неизвестное до тех пор вещество со свойствами слабой кислоты. Позднее химик Левин установил, что в состав этой кислоты входит углевод дезоксирибоза (см. приложения), отчего она и была названа дезоксирибонуклеиновой (ДНК). В 1920 г. тот же Левин идентифицировал в составе ДНК четыре азотистых основания: аденин (А), гуанин (Г), цитозин (Ц) и тимидин (Т) (см. приложения). Таким образом, уже в 20‑х годах XX в. ученые знали, из чего состоит ДНК. Эти сведения были существенно дополнены в 1950 г. биохимиком Чаргафом, обнаружившим, что в молекуле ДНК количество А равно количеству Т, а количество Г равно количеству Ц.
Однако, что касается роли ДНК в хранении и передаче наследственной информации, то долгое время по этому поводу имелись только догадки. В 1944 г. микробиологи Эвери, Маккарти и Маклеод впервые передали от одного микроба к другому определенные свойства с помощью ДНК.
А 28 февраля 1953 г. два молодых ученых из Кембриджского университета Джеймс Уотсон и Френсис Крик сообщили о своем открытии структуры молекулы ДНК.
Открытие ДНК
В 1951 году, когда Джеймс Уотсон и Фрэнсис Крик начали совместную работу, Уотсон в свои 23 года уже защитил диссертацию, в то время как Крик был 35-летним аспирантом. Но их объединил интерес к вопросу о хранении генетической информации в молекулярной форме. Уотсон и Крик много размышляли о ДНК, о возможной подходящей модели структуры. Ключевую роль в разгадке структуры сыграла фотография, полученная Морисом Уилкинсом, Розалинд Франклин и аспирантом Раймондом Гослингом.
В ноябре 1951 года Уилкинс поделился своими данными с Уотсоном и Криком. Александр Стокс , другой специалист в теории дифракции, и Уилкинс пришли к выводу, что данные по дифракции рентгеновских лучей для ДНК указывают на то, что молекула имеет спиральную структуру, но Франклин не соглашалась с этим выводом. По этим результатам в 1951 году Дж. Уотсон и Ф. Крик предложили и опубликовали модель структуры ДНК, которая оказалась ошибочной. Ученые хорошо понимали, что их главный конкурент, Лайнус Полинг, может опередить их открытие в любой момент. Их настораживал успех Полинга в открытии альфа-спирали белка, они опасались, что Полинг может также стать первым, кто определит правильную структуру ДНК.
Неоспоримую роль для построения модели ДНК было понимание Розалинд Франклин базовых законов химии: гидрофильные фосфат-содержащие группы нуклеотидной цепи ДНК должны быть расположены так, чтобы взаимодействовать с молекулами воды (то есть находиться на внешней стороне молекулы), в то время как гидрофобные азотистые основания должны быть обращены внутрь. Франклин поделилась своими соображениями с Уотсоном и Криком, тем самым она указала на главную ошибку предложенной ими модели в 1951 году.
Другими подсказками к открытию правильной структуры ДНК были так называемые соотношения Чаргаффа — экспериментально определенные соотношения нуклеотидных субъединиц ДНК: количество гуанина равно количеству цитозина и количество аденина равно количеству тимина. Приезд Эрвина Чаргаффа в Англию в 1952 году лишний раз подчеркнул этот важный факт для Уотсона и Крика. Для определения структуры ДНК эти соотношения не находили никакого применения до тех пор, пока Уотсон, занимаясь построением структурных моделей, понял, что А—T и C—G пары структурно похожи. В частности, длины этих пар оснований одинаковы. Чаргафф также заметил Уотсону, что в водной среде клетки преобладающими таутомерами пиримидиновых оснований (C и Т) будут амино- и кето- таутомеры цитозина и тимина, а не иминные и енольные формы, как изначально считали Крик и Уотсон. Они также консультировались с Джерри Донохью, который подтвердил наиболее вероятные структуры нуклеотидных оснований. Азотистые основания удерживались вместе водородными связями — теми же нековалентными взаимодействиями, которые стабилизировали белок α-спирали. Правильные структуры были важны для определения правильного расположения водородных связей. После открытия водородного связывания между парами A:T и C:G, Уотсон и Крик вскоре расставили цепи нуклеотидов антипараллельно в виде двойной спирали, азотистые основания были обращены внутрь спирали и соединялись между собой водородными связями. Тем самым был наглядно представлен способ распаковки двух комплементарных цепей (разрыв водородных связей) в репликации. Возможность репликации была последним требованием к модели генетической молекулы. Впоследствии Крик отметил, что без сотрудничества с Уотсоном он не смог бы найти правильную структуру молекулы ДНК самостоятельно.
Крик предварительно пытался поставить несколько экспериментов для установления закономерностей связывания нуклеотидов между собой, но он был больше биологом-теоретиком, чем экспериментатором. Крик начал думать о взаимодействиях между основаниями. В начале 1952 года он попросил Джона Гриффита вычислить силы притяжения между основаниями ДНК из химических принципов и законов квантовой механики. Наилучшие результаты Гриффит получил при рассмотрении взаимодействий пар А—T и G—C. В то время Крик не знал о правилах Чаргаффа, но эти немногочисленные расчеты навели учёного на мысль о возможной комплементарности азотистых оснований. Окончательные правильные соотношения (A-T, G-C) были получены Уотсоном. Он нарезал из картона детали, моделирующие молекулы пуринов и пиримидинов и стал раскладывать вырезки на столе подобно тому, как Лайнус Полинг открыл альфа-спираль несколькими годами раньше. Уотсон и Крик смогли открыть двойную спираль ДНК благодаря их готовности совмещать теорию, моделирование и экспериментальные результаты (хотя эти результаты были получены другими учеными).
Структура двойной спирали ДНК, предложенная Уотсоном и Криком, основывалась на связях «Уотсона — Крика» между четырьмя основаниями, которые наиболее часто встречаются в ДНК (A, C, T, G) и РНК (A, C, U, G). Однако более поздние исследования показали, что для трехцепочечных и четырёхцепочечных и других более сложных молекулярных структур ДНК необходим альтернативный вариант связывания — Хугстиновские пары. Кроме того, биохимики-синтетики начали поиск альтернативных синтетических ДНК, построенных из азотистых оснований, отличных от аденина, тимина, цитозина, гуанина. Также предпринимались попытки создать синтетический кодон (последовательность из трех нуклеотидов, точно определяющая одну аминокислоту), синтетические эндонуклеазы, синтетические белки и синтетические цинковые пальцы. В синтетической ДНК вместо 43 кодонов (из 4 азотистых оснований у природной ДНК) можно получить уже n3 кодонов (из n-азотистых оснований у синтетической ДНК). Новые кодоны могут участвовать в формировании новых аминокислот, которые, в свою очередь, сформируют новые белки.
Датой открытия двойной спирали ДНК считается 28 февраля 1953 года. Статья Уотсона и Крика была опубликована в журнале «Nature» 25 апреля. Её содержание было дублировано публичным докладом заведующего лабораторией, в которой работали Уотсон и Крик, Уильямом Брэггом, 14 мая. Уже 15 мая о нём была помещена заметка «Why You Are You. Nearer Secret of Life» в лондонской газете «News Chronicle». Victor K. McElheny в книге «Watson and DNA: Making a Scientific Revolution» указывает на короткую заметку в газете «The New York Times», состоящую всего из 6 абзацев с названием «Form of ‘Life Unit' in Cell Is Scanned», датированную 16 мая 1953 года. Заметка вышла только в ранних тиражах газеты, затем была заменена на другие новости. Затем, 12 июня 1953 года, в «The New York Times» вышла длинная статья об открытии структуры ДНК.
ДНК и её структура
Дезоксирибонуклеиновая кислота (ДНК) — макромолекула (одна из трёх основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК содержит информацию о структуре различных видов РНК и белков.
В клетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. Кроме того, одно- или двухцепочечные молекулы ДНК могут образовывать геном ДНК-содержащих вирусов.
С химической точки зрения ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы (фосфодиэфирные связи). В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула закручена по винтовой линии. В целом структура молекулы ДНК получила традиционное, но ошибочное название «двойной спирали», на самом же деле она является «двойным винтом». Винтовая линия может быть правой (A- и B-формы ДНК) или левой (Z-форма ДНК).
Дезоксирибонуклеиновая кислота (ДНК) представляет собой биополимер (полианион), мономером которого является нуклеотид.
Каждый нуклеотид состоит из остатка фосфорной кислоты, присоединённого дезоксирибозе, к которому также через гликозидную связь присоединено одно из четырёх азотистых оснований. Именно наличие характерного сахара и составляет одно из главных различий между ДНК и РНК, зафиксированное в названиях этих нуклеиновых кислот (в состав РНК входит сахар рибоза).
Исходя из структуры молекул, основания, входящие в состав нуклеотидов, разделяют на две группы: пурины (аденин [A] и гуанин [G]) образованы соединёнными пяти- и шестичленным гетероциклами; пиримидины (цитозин [C] и тимин [T]) — шестичленным гетероциклом.
Полимер ДНК обладает довольно сложной структурой. Нуклеотиды соединены между собой ковалентно в длинные полинуклеотидные цепи. Эти цепи в подавляющем большинстве случаев (кроме некоторых вирусов, обладающих одноцепочечными ДНК-геномами) попарно объединяются при помощи водородных связей во вторичную структуру, получившую название двойной спирали. Внутри одной цепи ДНК соседние нуклеотиды соединены фосфодиэфирными связями, которые формируются в результате взаимодействия между 3-гидроксильной (3—ОН) группой молекулы дезоксирибозы одного нуклеотида и 5-фосфатной группой (5—РО3) другого. Асимметричные концы цепи ДНК называются 3 (три прайм) и 5 (пять прайм). Полярность цепи играет важную роль при синтезе ДНК (удлинение цепи возможно только путём присоединения новых нуклеотидов к свободному 3-концу).
Как уже было сказано выше, у подавляющего большинства живых организмов ДНК состоит не из одной, а из двух полинуклеотидных цепей. Эти две длинные цепи закручены одна вокруг другой в виде двойной спирали, стабилизированной водородными связями, образующимися между обращёнными друг к другу азотистыми основаниями входящих в неё цепей. В природе эта спираль, чаще всего, правозакрученная. Направления от 3-конца к 5-концу в двух цепях, из которых состоит молекула ДНК, противоположны друг другу.
Ширина двойной спирали составляет от 22 до 24 А, или 2,2—2,4 нм, длина каждого нуклеотида 3,3 А (0,33 нм). Подобно тому, как в винтовой лестнице сбоку можно увидеть ступеньки, на двойной спирали ДНК в промежутках между фосфатным остовом молекулы можно видеть рёбра оснований, кольца которых расположены в плоскости, перпендикулярной по отношению к продольной оси макромолекулы.
В двойной спирали различают малую (12 А) и большую (22 А) бороздки. Белки, например, факторы транскрипции, которые присоединяются к определённым последовательностям в двухцепочечной ДНК, обычно взаимодействуют с краями оснований в большой бороздке, где те более доступны.
Если взяться за концы верёвки и начать скручивать их в разные стороны, она становится короче и на верёвке образуются «супервитки». Так же может быть суперскручена и ДНК. В обычном состоянии цепочка ДНК делает один оборот на каждые 10,459 основания, но в суперскрученном состоянии спираль может быть свёрнута туже или расплетена. Выделяют два типа суперскручивания: положительное — в направлении нормальных витков, при котором основания расположены ближе друг к другу; и отрицательное — в противоположном направлении. В природе молекулы ДНК обычно находятся в отрицательном суперскручивании, которое вносится ферментами — топоизомеразами. Эти ферменты удаляют дополнительное скручивание, возникающее в ДНК в результате транскрипции и репликации.
ПРИЛОЖЕНИЯ
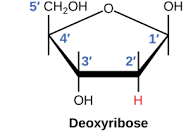 Дезоксирибоза
Дезоксирибоза
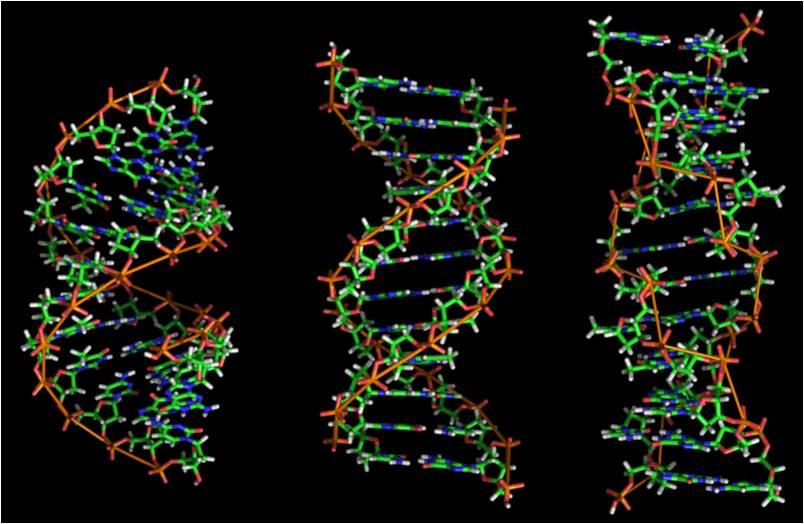
Модель строения молекулы ДНК
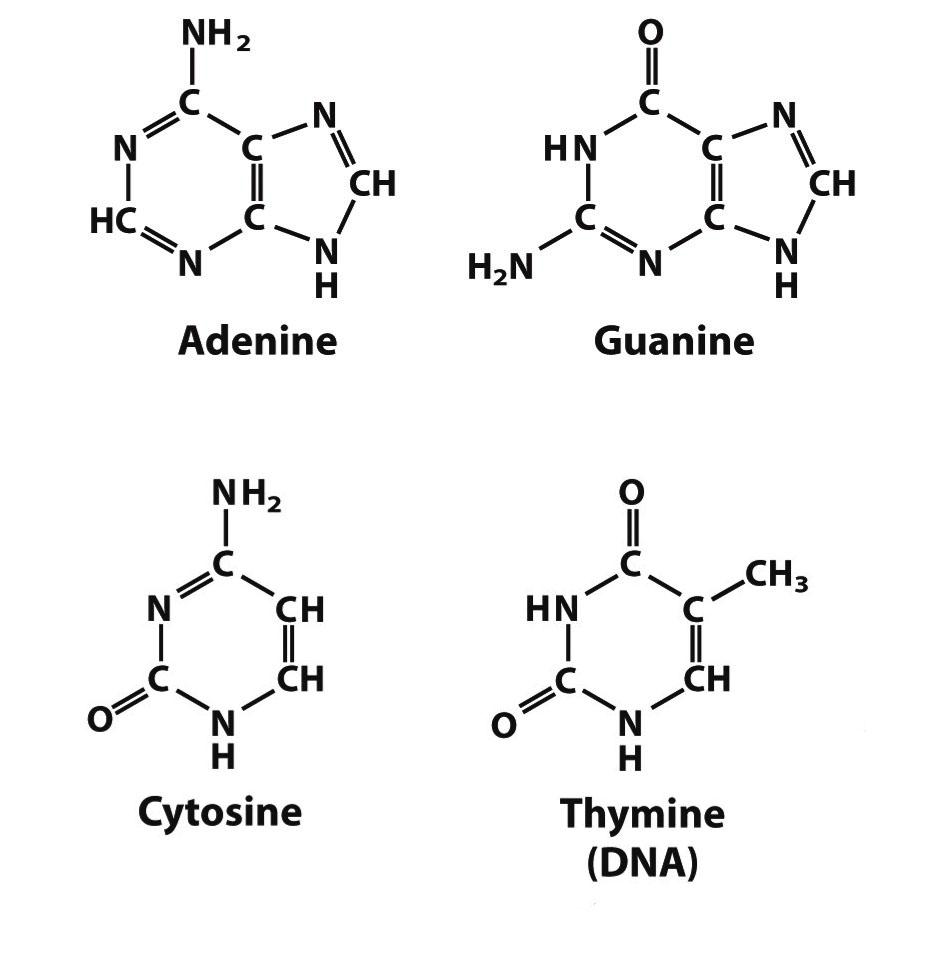
Азотистые основания


Дж. Уотсон Ф. Крик














Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!
Полезное для учителя

Реализация образовательных программ осуществляется с применением исключительно электронного обучения и ДОТ




Закрыть через 5 секунд