
Россия, Калининград


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Была в сети 06.06.2018 00:05

Мудрицкая Светлана Викторовна
заместитель директора, учитель биологии
76 лет
Местоположение
Специализация
Программа элективного курса "Путешествие в мир клетки" для 9-11 классов
Категория:
Биология
28.05.2018 20:22
Учебник:
Биология. Общая биология. Профильный уровень. 10 класс. Захаров В.Б. и др. М.: 2010. - 352 с.

Просмотр содержимого документа
«СЛОВАРЬ ЦИТОЛОГИЧЕСКИХ ТЕРМИНОВ»
Просмотр содержимого документа
«клетка модуль 1»
Муниципальное общеобразовательное учреждение средняя общеобразовательная школа №50
Учебное пособие к программе факультативного курса для 10-11 классов
СоставлЕНО на основе материалов
сайта:http://www.RUSOLIMP.ru/
г. Калининград
2009 г.
Введение
Учебное пособие блочно - модульного типа «Мир клетки» подготовлено для проведения факультативного курса для учащихся 10-11 классов. Необходимость введения факультативного курса определяется в первую очередь тем, что за последние двадцать лет произошло значительное развитие биологической науки: гигантский шаг вперед сделала молекулярная биология и молекулярная генетика, клеточная и молекулярная инженерия. За эти годы накопилось множество новых сведений о хроматине и хромосомах, о мембранах клетки и цитоскелетных структурах и т.д. В школьных же учебниках определенный объем теоретического материала в области цитологии устарел, зачастую используется старая номенклатура. В учебном пособии «Мир клетки» изложены основные вопросы о морфофункциональных особенностях прокариотической и эукариотической клеток на микроскопическом и ультрамикроскопическом уровне.
Освещены такие вопросы, как основные положения клеточной теории, формы клеточной гибели (некроз и апоптоз), поскольку эти вопросы включены в стандарт общего образования в области биологии, а в школьных учебниках они не рассматриваются или рассматриваются недостаточно подробно.
Изучение клеток бактерий, растений, грибов, животных осуществлялось в 6 – 9 классах, и элективный курс направлен на углубление, систематизацию и обобщение знаний, таким образом, элективный курс является интегративным. И, наконец, целью курса является формирование умений отличать типы клеток и их органеллы на микропрепаратах, что определено образовательным стандартом и не всегда представляется возможным из-за лимита времени на уроке, а главное - материально-технических возможностей.
В учебное пособие введены сведения о последних достижениях науки. Ознакомление школьников с новинками науки крайне необходимо для понимания, чем живет наука в данный момент, какие «горячие точки» в ней привлекают внимание исследователей.
Одной из особенностей пособия является то, что оно построено в структуре учебного модуля и кроме информационного блока, снабженного большим количеством иллюстраций, содержит рекомендации для самостоятельного освоения учебного материала, задания в тестовой форме для самоконтроля, задания для практической работы, ситуационные задачи и контрольные задания повышенного уровня сложности.
Предисловие
Вы знаете, что изучение мира живого - занятие столь же древнее, как и возраст человечества, поскольку изучение и сравнение удивительно разнообразных живых форм всегда было связано с самыми насущными потребностями человеческой жизни. Наши предки отлично разбирались в различиях между летучей мышью и птицей задолго до того, как они стали подозревать о существовании сходства между тканями гриба и тканями человеческого организма.
Данное учебное пособие посвящено изучению основных вопросов биологии клетки и построено по модульному типу.
Понятие «модуль» - блок чаще всего используется в точных и технических науках, и обозначает структурно-функциональный. Все чаще принцип модульной организации обучения используется для самостоятельного освоения учебной дисциплины или элективного курса.
Содержания пособия разбито на четыре блока - раздела. По оглавлению легко найти содержание данных блоков.
Задания вводного теста в начале пособия помогут Вам определить базовый уровень подготовки по вопросам цитологии, поскольку Вы изучали строение клеток различных организмов: бактерий, грибов, растений и животных в шестом, седьмом, восьмом и девятом классах.
В начале каждого раздела даны целевые установки, то есть информация о том, что Вы узнаете, чему научитесь при изучении данного раздела, причем, многие пункты целевой программы взяты непосредственно из федерального компонента государственного стандарта общего образования.
За целевыми установками следует информационный блок, содержащий собственно учебный материал. Учебный материал иллюстрирован схемами, рисунками, микрофотографиями; весь иллюстративный материал поможет Вам наиболее полно осмыслить изучаемый материал. Поэтому внимательно читайте подписи и объяснения к иллюстрациям. Все термины и незнакомые слова выделены жирным курсивом и имеют объяснения, как в тексте, так и в словаре, который завершает учебное пособие.
После информационного блока следуют рекомендации для самоподготовки. Эти рекомендации включают вопросы для обсуждения и осмысления учебного материала, задания для его систематизации и обобщения.
Каждый раздел учебного пособия содержит комплект заданий в тестовой форме, при выполнении которого Вы сможете оценить уровень усвоения учебной информации.
Практические занятия, ситуационные задачи, завершающие каждый раздел направлены на формирование необходимых умений и компетенций.
Учебный модуль содержит список использованной литературы и электронных информационных средств.
Мы надеемся, что данное пособие позволит Вам не только успешно подготовиться к вступительным экзаменам институты, к участию в олимпиадах, но и научит самостоятельно пополнять и совершенствовать свои знания и умения, а также выработает у Вас способность уверенно применять свои знания, навыки и умения в конкретных ситуациях.
Вводный тест
Дополните:
Какие жизненные процессы присущи растительной клетке ______________________
Оболочка растительной клетки обладает большой прочностью потому, что она __________.
Какой газ растительная клетка поглощает при дыхании ____________________.
Как называются тельца, передающие наследственные признаки от клетки к клетке ________.
Споры бактерий служат для _______________________ .
Грибы отличаются от растений тем, что их клеточная стенка содержит _________________ .
Выберите номер правильного ответа.
7. Ядро в клетке
1) содержит запас питательных веществ;
2) осуществляет связь между органоидами и частями клетки;
3) способствует поступлению веществ в клетку;
4) обеспечивает сходство материнской клетки с дочерними.
8. В основе роста растительного и животного организма лежит процесс
1) деления клеток;
2) оплодотворения;
3) обмена веществ;
4) пищеварения.
Пластиды зеленого цвета называются
хромопласты;
хлоропласты;
лейкопласты.
Зрелая растительная клетка отличается от молодой тем, что
вакуоль крупная, ядро находится в центре;
вакуоли мелкие, ядро находится в центре;
вакуоли мелкие, ядро расположено на периферии;
вакуоль крупная, ядро - на периферии.
11.Органеллами движения у амебы являются
жгутики;
реснички;
ложноножки.
12. Элементарной структурно-функциональной единицей всего живого является
зигота
росток
семя
клетка
13. Наследственные свойства клетки несет
ядерный сок
хромосома
ядерная оболочка
ядрышко
Выберите номера всех правильных ответов
12. Клетка бактерий отличается от клетки животного
наличием наружной мембраны
отсутствием ядра
отсутствием цитоплазмы
наличием плотной оболочки
отсутствием митохондрий
содержанием органических веществ
(Проверьте правильность выполнения тестовых заданий по эталону на странице….)
Раздел 1. Предмет цитологии. Методы изучения клетки
При изучении этого раздела Вы узнаете:
что изучает цитология;
основные этапы становления цитологии как науки;
сущность положений клеточной теории;
сущность традиционных и современных методов изучения морфофизиологии клетки
ИНФОРМАЦИОННЫЙ БЛОК
Цитология относится к фундаментальным разделам биологии, т.к. она исследует и описывает единственную единицу всего живого на Земле – клетку. Клетка не только основная структурная, но и функциональная, и генетическая единица организации живого, элементарная живая система. Клетка может существовать как отдельный организм (бактерии, простейшие, некоторые водоросли и грибы) или в составе тканей многоклеточных животных, растений, грибов.
. Основные этапы становления цитологии как науки
Познание клетки имеет важнейшее значение для развития множества других биологических наук, таких как физиология, генетика, молекулярная биология и др., поскольку дает им материал для изучения отдельных свойств именно клеток, а все физиологические процессы организмов имеют клеточную основу. Огромное значение современная цитология имеет для медицины, так как любые заболевания человеческого организма своей основой имеют патологию конкретных клеток или их групп, что важно для понимания развития болезни, для ее диагностики и для выбора методов лечения и профилактики заболевания.
Во второй половине XX века цитология из описательно-морфологической превратилась в экспериментальную науку, которая ставит перед собой задачи изучения физиологии клетки на ультраструктурном уровне, что связано с достижениями биохимии, биофизики, молекулярной биологии и генетики. Это послужило основанием для углубленного изучения морфофизиологии клетки с позиций данных наук, что обеспечило появление синтетической науки о клетке, которую чаще называют клеточной биологией. В этой науке сочетаются как морфологические, так и молекулярно-биологические подходы, что позволяет в настоящее время считать термины цитология и биология клетки совпадающими, т.к. предметом их изучения является клетка, имеющая свои собственные закономерности организации и функционирования.
Чуть более 300 лет отделяют нас от того времени, когда была впервые увидена клетка. Мир клеток невидим невооруженным глазом, он оставался полностью неизведанным до середины XVII столетия.
Одним из первых создателей микроскопа был англичанин Роберт Гук – физик, метеоролог, биолог, инженер, архитектор. В 1665 году он опубликовал альбом рисунков под названием « Микрография», изображающих его наблюдения под микроскопом. Среди рисунков был тонкий срез пробковой ткани дерева, структура которого напоминала пчелиные соты. Роберт Гук дал им название ячейки или клетки (от греч. «kytos» - ячейка, клетка). Это слово закрепилось для обозначения живых клеток, и вошло в название науки, их изучающей.
Микрография», изображающих его наблюдения под микроскопом. Среди рисунков был тонкий срез пробковой ткани дерева, структура которого напоминала пчелиные соты. Роберт Гук дал им название ячейки или клетки (от греч. «kytos» - ячейка, клетка). Это слово закрепилось для обозначения живых клеток, и вошло в название науки, их изучающей.
Работы Р. Гука вызвали интерес к дальнейшим микроскопическим исследованиям организмов. Современник Гука Антони ван Левенгук создал более двухсот микроскопов особой конструкции, позволяющих давать увеличение в 270 раз. Он впервые увидел и описал (1680) клетки животных (эритроциты и сперматозоиды), а так же удивительный мир бактерий - «анимакулюсов».
М . Мальпиги и Н. Грю на заседании Королевского ботанического общества (1671) доложили о том, что растительные клетки не однородны, среди них есть пузырьки и волокнистые структуры. Позднее клетки животных описал Ф.Фонтана (1781). Однако возможности светового микроскопа в XVII-XVIII веках были ограничены.
. Мальпиги и Н. Грю на заседании Королевского ботанического общества (1671) доложили о том, что растительные клетки не однородны, среди них есть пузырьки и волокнистые структуры. Позднее клетки животных описал Ф.Фонтана (1781). Однако возможности светового микроскопа в XVII-XVIII веках были ограничены.
Накопление материала о клеточном строении растений и животных, о структуре самих клеток шло медленно, и многочисленные исследования не привели в то время к пониманию универсальности клеточного строения, к четким представлениям о том, что представляет собой клетка.
Прогресс в изучении микроанатомии клетки связан с развитием микроскопической техники в XIX веке. К этому времени изменились представления о строении клетки: главным в организации клетки стала считаться не клеточная стенка, а собственно ее содержимое - протоплазма (Я. Пуркинье, 1830). Позднее в 1833 М. Брауном в протоплазме был открыт постоянный компонент клетки - ядро.
Длительное и пристальное изучение клетки привело к формулированию важного теоретического обобщения, так называемой клеточной теории, имеющей огромное общебиологическое значение.
1.2. Основные положения клеточной теории
Клеточная теория – это обобщенные представления о строении клеток как единиц живого, об их размножении и роли в формировании многоклеточных организмов.
Формулированию отдельных положений клеточной теории предшествовал довольно длительный период накопления наблюдений над строением различных одноклеточных и многоклеточных организмов растений и животных.
 Исследования в области микроскопии позволили в тридцатые годы XIX века ботанику Матиасу Шлейдену (1838) и зоологу Теодору Шванну («Микроскопические исследования о соответствии в структуре и росте животных и растений»1839) сделать фундаментальные обобщения о клеточной организации живого и сформулировать основные положения клеточной теории. «Заслуга Т. Шванна заключалась не в том, что он открыл клетки как таковые, а в том, что он научил исследователей понимать их значение» Вальдейер, 1909.
Исследования в области микроскопии позволили в тридцатые годы XIX века ботанику Матиасу Шлейдену (1838) и зоологу Теодору Шванну («Микроскопические исследования о соответствии в структуре и росте животных и растений»1839) сделать фундаментальные обобщения о клеточной организации живого и сформулировать основные положения клеточной теории. «Заслуга Т. Шванна заключалась не в том, что он открыл клетки как таковые, а в том, что он научил исследователей понимать их значение» Вальдейер, 1909.
Дальнейшее развитие клеточная теория получила в работах патолога Рудольфа Вирхова, который в 1858 году провозгласил: Omnis cellula e cellula (Всякая клетка происходит из другой клетки). Это утверждение стало четвертым положением клеточной теории.
Создание клеточной теории стало важнейшим событием в биологии, одним из решающих доказательств единства всей живой природы. Клеточная теория оказала значительное влияние на развитие биологии, послужила главным фундаментом для развития таких дисциплин, как эмбриология, гистология и физиология. Она дала основы для понимания жизни и механизмов индивидуального развития, для объяснения родственной взаимосвязи организмов.
Основные положения клеточной теории сохранили свое значение и на сегодняшний день, хотя более чем за сто пятьдесят лет были получены новые сведения о структуре, жизнедеятельности и развитии клеток.
Клетка – элементарная единица живого; вне клетки нет жизни.
Клетка – единая, целостная система, состоящая из множества закономерно связанных друг с другом элементов – органелл или органоидов.
Клетки сходны (гомологичны) по строению и по основным свойствам.
Клетка происходит от клетки. Клетки увеличиваются в числе путем деления исходной клетки после удвоения ее генетического материала (ДНК).
Многоклеточный организм представляет собой сложный ансамбль из множества клеток, объединенных в системы тканей и органов, связанных друг с другом с помощью гуморальных и нервных факторов.
Клетки многоклеточных организмов тотипотентны, т.е. обладают генетическими потенциями всех клеток данного организма, равнозначны по генетической информации, но отличаются друг от друга разной экспрессией (работой) различных генов, что приводит к их морфологическому и функциональному разнообразию – дифференцировке.
Давайте подробнее рассмотрим сущность основных положений клеточной теории.
1. Клетка – элементарная единица живого
Представление о клетке как о единице живого было дано еще в работах Т. Шванна. Р. Вирхов также считал, что каждая клетка несет в себе полную характеристику жизни: «Клетка есть последний морфологический элемент всех живых тел, и мы не имеем права искать настоящей жизнедеятельности вне ее» (1858). Современная наука полностью доказала это положение. В популярной литературе клетку часто называют «атомом жизни», «квантом жизни», подчеркивая тем самым, что клетка – это наименьшая единица живого, вне которой нет жизни.
Такая общая характеристика клетки должна в свою очередь опираться на определение живого. М.В. Волькенштейн (1965) дает следующее определение жизни: «Живые организмы представляют собой открытые, саморегулирующиеся и самовоспроизводящиеся системы, важнейшими функционирующими веществами которых являются белки и нуклеиновые кислоты». Живому свойствен ряд совокупных признаков: способность к воспроизведению, использование и трансформация энергии, обмен веществ (метаболизм), чувствительность и изменчивость. И такую совокупность признаков можно обнаружить на клеточном уровне. Среди живых организмов встречаются два типа организации клеток. К наиболее простому типу строения можно отнести клетки бактерий и сине-зеленых водорослей – цианобактерий, архибактерий к более высокоорганизованному - клетки всех остальных живых существ, начиная от одноклеточных животных и заканчивая человеком. Принято называть клетки бактерий и сине-зеленых водорослей прокариотическими (доядерными клетками), а клетки всех остальных представителей живого – эукариотическими (собственно ядерными), так как у последних обязательной структурой служит клеточное ядро, отделенное от цитоплазмы ядерной оболочкой.
Содержимое прокариотической клетки одето плазматической мембраной. Обычно снаружи от плазматической мембраны расположена клеточная стенка. У прокариотических клеток нет морфологически выраженного ядра, но присутствует нуклеоид - молекула ДНК с белком.
В цитоплазме прокариотических клеток располагаются многочисленные рибосомы, цитоплазматические же мембраны обычно выражены не так сильно, как у эукариотических клеток. Обычно все внутриклеточные мембранные системы прокариот (например, мезосомы) развиваются за счет наружной плазматической мембраны.
У эукариотических клеток кроме ядра в цитоплазме присутствуют органеллы, выполняющие определенные функции. К числу органелл относят мембранные и немембраные структуры. Эукариотические клетки, как правило, намного крупнее прокариотических. Так, палочковидные бактерии имеют длину до 5 мкм, а толщину около 1 мкм, в то время как эукариотические клетки могут достигать десятков мкм.
Несмотря на четкие морфологические отличия, и прокариотические и эукариотические клетки имеют много общего, что и позволяет отнести их к одной, клеточной, системе организации живого. В клетках прокариот и эукариот сходно происходят процессы активного межмембранного транспорта, синтез белка, РНК и ДНК, похожи и биоэнергетические процессы.
Таким образом, клетка – это ограниченная активной мембраной, упорядоченная структурированная система биополимеров (белков, нуклеиновых кислот и др.) и их макромолекулярных комплексов, участвующих в единой совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
Одно время первый постулат клеточной теории подвергался многочисленным нападкам и критике. Некоторые авторы указывали, что в многоклеточных организмах, особенно у животных, кроме клеток существуют и межклеточные, промежуточные вещества, которые тоже, казалось, обладали свойствами живого. В настоящее время известно, что компоненты межклеточного вещества (основное вещество и волокна) представляют собой не самостоятельные образования, а производные определенных групп клеток.
Другие возражения касались того, что часто у животных кроме отдельных клеток встречаются надклеточные структуры: так называемые симпласты и синцитии (соклетия). По морфологическому описанию это крупные цитоплазматические многоядерные образования. Примерами симпластов могут служить мышечные волокна позвоночных, а синцитиев - эпидермис у ленточных червей, а также плазмодии у низших грибов миксомицетов. В отличие от симпластов синцитии возникают вторично за счет слияния отдельных клеток или же в результате деления одних ядер без разделения цитоплазмы.
2. Клетка – единая система соподчиненных функциональных единиц
Изучая клетки растений, животных, человека, мы знаем о сложности строения этой «единицы», которая содержит в себе различные внутриклеточные структуры, выполняющие разнообразные функции. Формально любую клетку можно «разложить» на ряд как бы независимых структурных и функциональных компонентов, выполняющих свои специфические функции. Так, например, эукариотические клетки принято разделять на ядра и цитоплазму. В цитоплазме, в свою очередь, выделяют гиалоплазму или основную плазму клетки (цитозоль – растворимый компонент цитоплазмы по терминологии биохимиков), а также целый ряд структур – органелл, выполняющих свои отдельные специфические функции. Выделяют мембранные органеллы. К ним относятся: одномембранные (эндоплазматическая сеть, комплекс Гольджи, эндо- и экзоцитозные вакуоли, лизосомы, пероксисомы) и двумембранные (митохондрии и пластиды). К немембранным органеллам нужно отнести рибосомы и систему цитоскелетных фибрилл. Кроме того, вся поверхность клетки покрыта цитоплазматической мембраной, тесно функционально связанной как с вакуолярной системой, с элементами цитоскелета, так и с гиалоплазмой.
Каждая из перечисленных структур представляет собой подсистему функционирования клетки. Так клеточное ядро является системой хранения, воспроизведения и реализации генетической информации. Гиалоплазма - система основного промежуточного обмена; рибосомы – элементарные «клеточные машины» синтеза белка; цитоскелет - опорно-двигательная система клетки; вакуолярная система - система синтеза и внутриклеточного транспорта белковых биополимеров и образование многих клеточных мембран; митохондрии - органеллы энергообеспечения клетки за счет синтеза АТФ, пластиды растительных клеток - система синтеза АТФ и фотосинтеза, плазматическая мембрана - барьерно-рецепторно-транспортная система клетки. Все эти подсистемы клетки находятся во взаимозависимости. Так, например, нарушение функций ядра сразу сказывается на синтезе клеточных белков, нарушение работы митохондрий прекращает все синтетические и обменные процессы в клетке, разрушение элементов цитоскелета прекращает внутриклеточный транспорт и т.д. Как в часовом механизме, повреждение любой его части приводит к остановке всей системы в целом.
3. Гомологичность клеток
Термин гомологичность означает сходство по коренным свойствам и отличие по второстепенным. При изучении клеток растений и животных вы видели разительное сходство не только в микроскопическом строении этих клеток, но и в деталях строения их отдельных компонентов. Таким образом, у эукариот и прокариот клетки отделены друг от друга или от внешней среды активной плазматической мембраной, которая может принимать участие в выделении веществ из клетки и построении внеклеточных структур. У всех эукариотических клеток от низших грибов до позвоночных всегда имеется ядро, сходное по построению у разных организмов. Строение и функции внутриклеточных структур также в принципе определяется гомологичностью общеклеточных функций (синтез нуклеиновых кислот и белков, процессы энергетического обмена).
Одновременно мы видим и разнообразие клеток даже в пределах одного многоклеточного организма. Гистологи выделяют в организме человека более 200 типов клеток. Например, по форме мало похожи друг на друга такие клетки, как мышечная и нервная. В мышечной клетке кроме общеклеточных структур (мембранные системы ЭПС, аппарат Гольджи, рибосомы и др.) находятся миофиломенты, выполняющие сократительную функцию. В нервной клетке выделяют тигроид - мощно развитую эндоплазматическую сеть, большое количество нейротрубочек и нейрофиламентов, а также межклеточные контакты - синапсы. Вся совокупность этих отличительных черт нервной клетки связана с ее специализацией – передачей нервного импульса. Структурное разнообразие клеток многоклеточного организма можно объяснить отличием их специальных функций, осуществляющихся данной клеткой. Таким образом, гомологичность в строении клеток определяется сходством общеклеточных функций, направленных на поддержание жизни самих клеток и на их размножение. Разнообразие же в строении клеток многоклеточных – результат функциональной специализации.
4. Клетка от клетки
Формулировка положения «Всякая клетка от клетки» (Omnis cellula e cellula) связана с именем знаменитого ученого Р. Вирхова, который как противник идеи о самозарождении жизни, настаивал на «преемственном размножении клеток». Сегодня сформулированное Р. Вирховым определение можно считать биологическим законом. Размножение клеток, прокариотических и эукариотических, происходит только путем деления исходной клетки, которому предшествует воспроизведение ее генетического материала (редупликация ДНК).
У эукариотических клеток единственно полноценным способом деления является митоз (или мейоз при образовании половых клеток). При этом образуется специальный аппарат клеточного деления – клеточное веретено, с помощью которого равномерно и точно по двум дочерним клеткам распределяются хромосомы. Этот тип деления наблюдается у растительных и животных клеток. Прокариотические клетки, делящиеся так называемым бинарным образом, также используют специальный аппарат разделения клеток, значительно напоминающий митотический способ деления эукариот.
5. Клетки и многоклеточный организм
Теодор Шванн представлял себе многогранную деятельность организма как сумму жизнедеятельности отдельных клеток. Это представление было в свое время принято и расширено Р. Вирховым и получило название теории «клеточного государства». Вирхов писал: «…всякое тело, сколько-нибудь значительного объема, представляет устройство, подобное общественному, где множество отдельных существований поставлено в зависимость друг от друга, но так, однако же, что каждое из них имеет свою собственную деятельность, и если побуждение к этой деятельности оно и получает от других частей, зато самою работу свою оно совершает собственными силами» (1859).
Действительно, какую бы сторону деятельности целого организма мы ни брали, будь то реакция на раздражение или движение, иммунные реакции, выделение и многое другое, каждая из них осуществляется специализированными клетками. Клетка – это единица функционирования в многоклеточном организме. Но клетки объединены в функциональные системы, в ткани и органы, которые находятся во взаимной связи друг с другом. Многоклеточные организмы представляют собой сложные ансамбли клеток, объединенные в системы тканей и органов, подчиненные и связанные межклеточными, гуморальными и нервными формами регуляции. Вот почему мы говорим об организме как о целом.
Таким образом можно сказать, что клетка в многоклеточном организме – это единица функции и развития. Кроме того, основой всех нормальных и патологических реакций организма является клетка. Например, когда в организм попадают чужеродные белки (бактериальные), развивается иммунная реакция. При этом в крови появляются белки-антитела, которые связываются с чужими белками и их обезвреживают. Эти антитела образуются особыми клетками - плазмоцитами. Но чтобы плазмоциты начали вырабатывать антитела, необходимо взаимодействие лимфоцитов и макрофагов.
Жизнь нового организма начинается с зиготы, получившейся в результате слияния женской половой клетки (ооцита) со спермием. Рост организма, увеличение его массы есть результат размножения клеток и результат выработки ими разнообразных продуктов (например, вещества кости или хряща).
И наконец, именно поражение клеток или изменение их свойств является основой для развития всех без исключения заболеваний. Данное положение было впервые сформулировано Р. Вирховым (1858) в его знаменитой книге «Клеточная патология». Классическим примером клеточной обусловленности развития болезни может служить сахарный диабет, широко распространенное заболевание современности. Его причина - недостаточность функционирования лишь одной группы клеток, так называемых В-клеток островков Лангерганса в поджелудочной железе. Эти клетки вырабатывают гормон инсулин, участвующий в регуляции сахарного обмена организма.
6. Тотипотентность клеток
Как же возникают разнообразные типы клеток в многоклеточных организмах? Современная биология на базе представлений эмбриологии, молекулярной биологии и генетики считает, что индивидуальное развитие от одной клетки до многоклеточного зрелого организма - это результат последовательного, избирательного включения работы разных генов. Это приводит к появлению клеток со специфическими для них структурами и особыми функциями, т.е. к процессу, называемому дифференцировкой.
Любая клетка многоклеточного организма обладает одинаковым набором генов и всеми возможными потенциями для их проявления, но в разных клетках одни и те же гены могут находиться или в активном или в репрессированном состоянии. Эти представления базируются на большом экспериментальном материале. Стало возможным вырастить зрелое растение из одной его соматической клетки. Общим законом для многоклеточных растительных и животных организмов является то, что, несмотря на структурные и функциональные различия клеток данного организма, в генетическом отношении, они однородны, тождественны и тотипотентны.
1.3. Методы клеточной биологии
Цитология возникла как ветвь микроанатомии, и поэтому одним из основных методов, который используют цитологи, - это метод световой микроскопии. Революционным моментом в развитии современной цитологии и биологии вообще было применение электронной микроскопии, открывшей необычайно широкие перспективы. С введением электронной микроскопии в ряде случаев уже трудно провести границу между собственно цитологией и биохимией, они объединяются на уровне макромолекулярного изучения объектов (например, микротрубочек, мембран, микрофиламентов и т.д.)..
1.3.1. Световая микроскопия
Световой микроскоп, главный прибор биологии, представляет собой оптическую систему, состоящую из конденсора, объектива и окуляра. Пучок света от источника освещения собирается в конденсоре и направляется на объект. Пройдя через объект, лучи света попадают в систему линз объектива; они строят первичное изображение, которое увеличивается с помощью линз окуляра. Главная оптическая часть микроскопа, определяющая его основные возможности - объектив. В современных микроскопах объективы сменные, что позволяет изучать клетки при разных увеличениях. Главной характеристикой микроскопа как оптической системы является разрешающая способность.
Н о все же в световом микроскопе можно видеть частицы меньшей величины, чем 0,2 мкм. Это метод «темного поля», или, как его называли раньше, метод «ультрамикроскопии». Суть его в том, что подобно пылинкам в луче света (эффект Тиндаля), в клетке при боковом освещении светятся мельчайшие частицы (меньше 0,2 мкм), отраженный свет от которых попадает в объектив микроскопа. Этот метод успешно применяется при изучении живых клеток.
о все же в световом микроскопе можно видеть частицы меньшей величины, чем 0,2 мкм. Это метод «темного поля», или, как его называли раньше, метод «ультрамикроскопии». Суть его в том, что подобно пылинкам в луче света (эффект Тиндаля), в клетке при боковом освещении светятся мельчайшие частицы (меньше 0,2 мкм), отраженный свет от которых попадает в объектив микроскопа. Этот метод успешно применяется при изучении живых клеток.
Е сли же необработанные живые или мертвые клетки рассматривать в проходящем свете, то в них различаются только крупные детали из-за того, что они обладают иным коэффициентом преломления и поглощения световых лучей, чем окружающая среда. Большая же часть клеточных компонентов мало отличается по этим свойствам как от среды (воды или тканевых растворов), так и друг от друга и поэтому мало заметны и не контрастны. Для их изучения применяют особые методы и приборы. Один из таких приемов – метод фазово-контрастной микроскопии, широко использующийся для наблюдений за живыми клетками. Он основан на том, что отдельные участки прозрачной в общем клетки хоть мало, но все же отличаются друг от друга по плотности и по светопреломлению. Проходя через них, свет изменяет свою фазу, однако такое изменение фазы световой волны наш глаз не улавливает, так как он чувствителен только к изменению интенсивности света. В фазово-контрастном микроскопе в объектив вмонтирована специальная пластинка, проходя через которую, луч света испытывает дополнительный сдвиг фазы колебаний. При построении изображения взаимодействуют уже лучи, находящиеся в одной фазе либо в противофазе, но обладающие разной амплитудой - тем самым создается светло-темное контрастное изображение объекта.
сли же необработанные живые или мертвые клетки рассматривать в проходящем свете, то в них различаются только крупные детали из-за того, что они обладают иным коэффициентом преломления и поглощения световых лучей, чем окружающая среда. Большая же часть клеточных компонентов мало отличается по этим свойствам как от среды (воды или тканевых растворов), так и друг от друга и поэтому мало заметны и не контрастны. Для их изучения применяют особые методы и приборы. Один из таких приемов – метод фазово-контрастной микроскопии, широко использующийся для наблюдений за живыми клетками. Он основан на том, что отдельные участки прозрачной в общем клетки хоть мало, но все же отличаются друг от друга по плотности и по светопреломлению. Проходя через них, свет изменяет свою фазу, однако такое изменение фазы световой волны наш глаз не улавливает, так как он чувствителен только к изменению интенсивности света. В фазово-контрастном микроскопе в объектив вмонтирована специальная пластинка, проходя через которую, луч света испытывает дополнительный сдвиг фазы колебаний. При построении изображения взаимодействуют уже лучи, находящиеся в одной фазе либо в противофазе, но обладающие разной амплитудой - тем самым создается светло-темное контрастное изображение объекта.
1.3.2. Витальное (прижизненное) изучение клеток
Световой микроскоп позволяет видеть живые клетки. Для кратковременного наблюдения клетки помещают просто в жидкую среду на предметное стекло; если нужно длительное наблюдение за клетками, то используются специальные камеры. Это или плоские флаконы с отверстиями, закрытыми тонкими стеклами, или же разборные плоские камеры. В качестве объектов можно использовать свободноживущие клетки простейших и других одноклеточных организмов, клетки крови или же разобщенные тканевые клетки многоклеточных организмов как животного, так и растительного происхождения.
Для изучения клеток органов и тканей животных используют метод клеточных культур. Более простой вариант этого метода заключается в том, что в камеру, наполненную питательной средой, помещают небольшой кусочек живой ткани. Через некоторое время на периферии такого кусочка начинается деление и рост клеток. В другом случае вырезанный кусочек ткани слегка обрабатывают раствором фермента трипсина, что приводит к полному разделению клеток друг от друга. Затем такую взвесь отмытых клеток помещают в сосуд с питательной средой, где они опускаются на дно, прикрепляются к стеклу и начинают размножаться, образуя сначала колонии, а затем - сплошной клеточный пласт. Так растут однослойные клеточные культуры, очень удобные для прижизненных наблюдений.
При культивировании клеток вне организма кроме смены среды важно поддерживать и необходимую температуру (20о для хладнокровных животных и 37о - для теплокровных). Обязательным условием культивирования клеток является соблюдение стерильности. Существует целый ряд длительно культивируемых клеток - это специальные клеточные штаммы, приспособившиеся десятилетиями к росту вне организма. Большей частью это клетки опухолевого происхождения или значительно измененные клетки, которые приобрели свойства опухолевых клеток. Наблюдения за живыми клетками обычно регистрируются в виде фотографий, сделанных с помощью специальных фотонасадок к микроскопу. Живые клетки можно снимать и на кинопленку. С развитием компьютерных технологий с помощью специальных телекамер стало возможно получать изображение клеток прямо на мониторе компьютера, записывать их в памяти компьютера, всячески обрабатывать и получать отпечатки на принтерах цветных или черно-белых.
При исследовании живых клеток используют методы микрохирургии - оперативного воздействия на клетки. С помощью прибора микроманипулятора клетки разрезают, извлекают из них части, вводят различные вещества и т.д. Микроманипулятор совмещается с обычным микроскопом, в который наблюдают за ходом операции. Микрохирургическими инструментами служат стеклянные крючки, иглы, капилляры, которые имеют микроскопические размеры.
При изучении живых клеток широко используют флуоресцирующие красители и метод флуоресцентной микроскопии. Суть его заключается в том, что целый ряд веществ обладает способностью светиться (флуоресцировать, люминесцировать) при поглощении ими световой энергии. Можно применять метод флуоресцентной микроскопии, добавляя живым клеткам флуоресцирующие вещества – флуорохромы, которые избирательно связываются с определенными структурами клетки, вызывая их люминесценцию.
В Рис. 6 Люминесцирующие бактерии Рис. 7 Срез ткани, флуорохромированный акридиновым оранжевым.

(иммунофлюоресценция).
Обычные методы световой микроскопии трудно использовать для воспроизведения трехмерной картины изучаемого объекта из-за небольшой глубины резкости микроскопа. Обычно клетки рассматриваются как оптические разрезы на данной глубине фокуса. Для того, чтобы получить полную трехмерную реконструкцию объекта используют специальный сканирующий световой микроскоп. С помощью этого прибора получают серии последовательных оптических срезов, взятых с различной глубины и изображения которых накапливаются в компьютере, и по специальной программе реконструируется трехмерное изображение объекта.
1.3.3 Изучение фиксированных клеток
Несмотря на важность и достаточную простоту витальных наблюдений, большая часть сведений о структуре и свойствах клеток получена на фиксированном материале. Если клетку повредить, она начинает претерпевать ряд изменений, а после смерти клетки в ней активируются аутолитические ферменты, что приводит к грубым изменениям клеточной структуры. Следовательно, первым этапом в обработке кусочков, вырезанных из различных органов и тканей для микроскопических исследований, является фиксация. Фиксация предпринимается для сохранения тканей и органов в таком виде, в каком они были в живом организме к моменту смерти, т.е. фиксация имеет целью закрепление тканевых структур. Вещества, применяемые при фиксации, называются фиксаторами. Часто для фиксации используются альдегиды и их смеси с другими веществами. В качестве фиксаторов применяют также спирты, вызывающие необратимую денатурацию белков, осаждение нуклеиновых кислот и полисахаридов. Осаждающим действием обладают также фиксаторы, содержащие сулему или пикриновую кислоту. После фиксации объекты в дальнейшем можно подвергать дополнительной обработке. Одной из главных таких обработок является окрашивание клеток. Именно дополнительное окрашивание клеток позволило выявить в них массу ультраструктурных деталей.
Стекла с фиксированными мазками одноклеточных организмов или с клетками культуры ткани можно непосредственно помещать в красители. Но для окрашивания клеток в составе органов необходимо получить их срезы. Изучают такие срезы и отдельных клеток.
Для этого после фиксации кусочки органов обезвоживают в спиртах возрастающей концентрации, затем спирт замещают ксилолом, а ксилол – парафином. Таким образом, фиксированная ткань оказывается заключенной в твердую массу парафина, которую можно нарезать.
С резы толщиной до 5-10 мкм получают на специальном приборе – микротоме. Такие срезы приклеиваются на предметное стекло. Затем парафин среза растворяется в ксилоле, ксилол удаляется спиртами, которые замещаются водой. Теперь срезы можно окрашивать водными растворами красителей. Для изготовления постоянных препаратов окрашенные срезы снова обезвоживают и заключают в канадский бальзам под покровным стеклом. Такие препараты можно длительно хранить.
резы толщиной до 5-10 мкм получают на специальном приборе – микротоме. Такие срезы приклеиваются на предметное стекло. Затем парафин среза растворяется в ксилоле, ксилол удаляется спиртами, которые замещаются водой. Теперь срезы можно окрашивать водными растворами красителей. Для изготовления постоянных препаратов окрашенные срезы снова обезвоживают и заключают в канадский бальзам под покровным стеклом. Такие препараты можно длительно хранить.
Для окраски фиксированных тканей и клеток применяют различные натуральные и синтетические красители. Натуральные красители (гематоксилин, кармин и др.) употребляют в сочетании с протравами (окислы различных металлов), с которыми они образуют комплексные соединения.
Красители подразделяют на кислые и основные. Основные красители представляют собой красящие основания или их соли. Такие красители образуют химические связи с кислотными группами в структурах клетки. Следовательно, участки клеток, богатые кислотными группами, которые свяжутся с основными красителями, будут базофильными.
К
ислотные красители связываются с основными (щелочными) структурами клетки, следовательно, эти структуры проявляют ацидофилию или оксифилию. Существует множество смесей таких красителей, которые одновременно могут окрашивать различные участки клеток в разные цвета и тем самым повышать контрастность клеточных и внеклеточных компонентов. Таким образом, используя всевозможные красители, исследователи не только добиваются четкости морфологической картины клетки, но получают некоторые сведения о химизме той или иной структуры.
Ряд методов, направленных на выявление специфических химических веществ в клетке, получил название гистохимических и цитохимических. Методов цитохимического анализа очень много. Примером такого рода цитохимических реакций может быть широко применяемая реакция на ДНК - реакция Фёльгена. Также специфически можно определить локализацию белков реакциями на отдельные аминокислоты (тирозин, триптофан, аргинин и др.). Липиды и жиры обнаруживают в клетках специальными красителями (например, судан черный), хорошо растворяющимися и аккумулирующимися в жировых включениях. Целая группа цитохимических реакций связана с обнаружением ферментов. Общий принцип этих реакций в том, что в микроскоп видны не сами белковые ферменты, а места их локализации, которые обнаруживаются по продуктам их ферментативной активности.
Для выяснения локализации мест синтеза биополимеров, для определения путей переноса веществ в клетке, для наблюдения за миграцией или свойствами отельных клеток широко используют метод радиоавтографии - регистрации веществ, меченных изотопами. При радиоавтографическом исследовании клеткам в среду вводится предшественник одного из макромолекулярных соединений (например, аминокислота или нуклеотид), один из атомов которого замещен радиоактивным изотопом. Например, вместо 12С введен атом 14С, вместо водорода – тритий 3Н и др. В процессе синтеза в биополимер включится и меченная молекула предшественника. Регистрировать ее место в клетке можно с помощью фотоэмульсии. Если клетки в пласте или на срезе покрыть фотоэмульсией, то через некоторое время в результате распада изотопа – частицы, разлетающиеся хаотично в разных направлениях, попадут в зону чувствительного фотослоя и активируют в нем зерна бромистого серебра. Метод радиоавтографии – один из основных методов, позволяющих изучать динамику синтетических процессов, сравнить их интенсивность в разных клетках на одном и том же препарате.
1.3.4 Электронная микроскопия
Р ассматривая характеристики светового микроскопа, можно убедиться, что единственным путем увеличения разрешения оптической системы будет использование источника освещения, испускающего волны с наименьшей длиной. Таким источником может быть раскаленная нить, которая в электрическом поле выбрасывает поток электронов. Последний можно фокусировать, пропуская через магнитное поле. Это послужило основой для создания электронного микроскопа, в котором уже сейчас достигнуто разрешение в 0,1 нм. По принципу конструкции электронный микроскоп очень сходен с оптическим: в нем есть источник освещения (катод электронной пушки), конденсорная система (конденсорная магнитная линза), объектив (объективная магнитная линза), окуляр (проекционные магнитные линзы), только вместо сетчатки глаза электроны попадают на люминесцирующий экран или на фотопластинку.
ассматривая характеристики светового микроскопа, можно убедиться, что единственным путем увеличения разрешения оптической системы будет использование источника освещения, испускающего волны с наименьшей длиной. Таким источником может быть раскаленная нить, которая в электрическом поле выбрасывает поток электронов. Последний можно фокусировать, пропуская через магнитное поле. Это послужило основой для создания электронного микроскопа, в котором уже сейчас достигнуто разрешение в 0,1 нм. По принципу конструкции электронный микроскоп очень сходен с оптическим: в нем есть источник освещения (катод электронной пушки), конденсорная система (конденсорная магнитная линза), объектив (объективная магнитная линза), окуляр (проекционные магнитные линзы), только вместо сетчатки глаза электроны попадают на люминесцирующий экран или на фотопластинку.
Основная часть такого микроскопа представляет собой полый цилиндр (колонка микроскопа), из которого откачан воздух для того, чтобы не было взаимодействия электронов с молекулами газов и окисления вольфрамовой нити накаливания в катоде электронной пушки. Между катодом и анодом подается высокое напряжение (от 50 до 200-5000 кВ), что служит причиной ускорения электронов. В центре анода есть отверстие, проходя через которое, электроны формируют пучок, идущий вниз по колонке микроскопа. Линзы электронного микроскопа представляют собой электромагниты, поле которых может изменять путь электронов (как стеклянные линзы изменяют путь фотонов). В конденсорной линзе пучок электронов фиксируется и попадает на объект, с которым электроны взаимодействуют, отклоняются, рассеиваются, поглощаются или проходят без изменения. Электроны, прошедшие через объект, фокусируются объективной линзой, которая формирует увеличенное первичное изображение объекта. Так же как в световом микроскопе, объективная линза определяет его основные показатели. Первичное изображение увеличивается проекционной линзой и проецируется на экран, покрытый люминесцентным слоем, светящимся при попадании на него электронов. Вместо светящегося экрана изображение можно поместить на фотопластинку и получить снимок. На экранах и фотопластинках электронных микроскопов получают увеличение до 50 000 раз, в дальнейшем при фотопечати можно получить еще 10-кратное увеличение, так что конечное увеличение, при котором максимально реализуется разрешение, достигает 106 раз.
Д ля изучения структуры различных мембранных компонентов клетки используется метод замораживания–скалывания. Он заключается в том, что объект сначала быстро замораживают жидким азотом, а затем при той же температуре переносят в специальную вакуумную установку. Там замороженный объект механическим способом скалывается охлажденным ножом. При этом обнажаются внутренние зоны замороженных клеток. В вакууме часть воды, перешедшей в стекловидную форму, возгоняется («травление»), а поверхность скола последовательно покрывается тонким слоем испаренного углерода, а затем металла. Таким образом с замороженного и сохраняющего прижизненную структуру материала получают реплику с его скола. Затем уже в условиях комнатной температуры ткань или клетки растворяют в кислотах, но пленка-реплика при этом остается цела, ее изучают в электронном микроскопе. Этот метод имеет два преимущества: изучают реплики со сколов нативных образцов и исследуют рельеф поверхности мембран клетки, что недостижимо другими методами. Оказалось, что и в этом случае общая организация клетки и ее компонентов сходна с тем, что мы видим при химической фиксации или при криотомии. Этот метод позволил увидеть, что как на поверхности, так и в толщине клеточных мембран располагаются глобулы интегральных белков, что мембраны не однородны по своей структуре.
ля изучения структуры различных мембранных компонентов клетки используется метод замораживания–скалывания. Он заключается в том, что объект сначала быстро замораживают жидким азотом, а затем при той же температуре переносят в специальную вакуумную установку. Там замороженный объект механическим способом скалывается охлажденным ножом. При этом обнажаются внутренние зоны замороженных клеток. В вакууме часть воды, перешедшей в стекловидную форму, возгоняется («травление»), а поверхность скола последовательно покрывается тонким слоем испаренного углерода, а затем металла. Таким образом с замороженного и сохраняющего прижизненную структуру материала получают реплику с его скола. Затем уже в условиях комнатной температуры ткань или клетки растворяют в кислотах, но пленка-реплика при этом остается цела, ее изучают в электронном микроскопе. Этот метод имеет два преимущества: изучают реплики со сколов нативных образцов и исследуют рельеф поверхности мембран клетки, что недостижимо другими методами. Оказалось, что и в этом случае общая организация клетки и ее компонентов сходна с тем, что мы видим при химической фиксации или при криотомии. Этот метод позволил увидеть, что как на поверхности, так и в толщине клеточных мембран располагаются глобулы интегральных белков, что мембраны не однородны по своей структуре.
Метод получения реплик с микрорельефа образца широко применяется при изучении фибриллярных компонентов клетки.
М етод сканирующей электронной микроскопии позволяет изучать трехмерную картину поверхности клетки. При сканирующей электронной микроскопии тонкий пучок электронов пробегает по поверхности объекта и полученная информация передается на электронно-лучевую трубку. Изображение может быть получено в отраженных или вторичных электронах. При этом методе фиксированный и специальным образом высушенный объект покрывается тонким слоем испаренного металла (чаще всего золота), отражаясь от которого электроны попадают в приемное устройство, передающее сигнал на электронно-лучевую трубку. Благодаря огромной глубине фокуса сканирующего микроскопа, которая значительно больше, чем у просвечивающего, получается почти трехмерное изображение исследуемой поверхности. Разрешающая способность этого типа приборов несколько ниже, чем у просвечивающих электронных микроскопов, но уже сейчас выпускаются приборы с разрешением 3-5 нм .
етод сканирующей электронной микроскопии позволяет изучать трехмерную картину поверхности клетки. При сканирующей электронной микроскопии тонкий пучок электронов пробегает по поверхности объекта и полученная информация передается на электронно-лучевую трубку. Изображение может быть получено в отраженных или вторичных электронах. При этом методе фиксированный и специальным образом высушенный объект покрывается тонким слоем испаренного металла (чаще всего золота), отражаясь от которого электроны попадают в приемное устройство, передающее сигнал на электронно-лучевую трубку. Благодаря огромной глубине фокуса сканирующего микроскопа, которая значительно больше, чем у просвечивающего, получается почти трехмерное изображение исследуемой поверхности. Разрешающая способность этого типа приборов несколько ниже, чем у просвечивающих электронных микроскопов, но уже сейчас выпускаются приборы с разрешением 3-5 нм .
1.3.5 Фракционирование клеток
В цитологии широко применяют различные методы биохимии, как аналитические, так и препаративные. В последнем случае можно получить в виде отдельных фракций разнообразные компоненты и изучать их химический состав, ультраструктуру и свойства. Так, в настоящее время в виде чистых фракций получают практически любые клеточные органеллы и структуры: ядра, ядрышки, хроматин, ядерные оболочки, плазматическую мембрану, вакуоли эндоплазматической сети, его рибосомы, рибосомы гиалоплазмы, комплекса Гольджи, митохондрии, их мембраны, пластиды, пероксисомы, микротрубочки и т.д. В последнее время получены чистые фракции центриолей и ядерных пор.
Получение клеточных фракций начинается с общего разрушения клетки. Затем из гомогенатов уже можно выделять фракции. Одним из основных способов выделения клеточных структур является центрифугирование. Его принцип применения в том, что время для осаждения частиц в гомогенате зависит от их размера и плотности: чем больше частица или чем она тяжелее, тем быстрее она осядет на дно пробирки. При центрифугировании раньше всего осядут ядра и не разрушенные клетки, затем осядут крупные частицы, состоящие из митохондрий, мелких пластид, пероксисом, лизосом.
Прежде чем их анализировать биохимическими способами полученные фракции, их необходимо проверить на чистоту с помощью электронного микроскопа.

1.3.5. Методы генной и клеточной инженерии
Большой вклад в биологию клетки вносят методы клеточной инженерии. Было найдено, что различные живые клетки могут сливаться друг с другом, если специальными способами обработать их плазматические мембраны. Так можно слить эритроцит курицы и лимфоцит человека. При этом получается двуядерная клетка, гетерокарион, в котором происходит активация ядра куриного эритроцита. Если гетерокарион образуется из близкородственных клеток (например, мыши и хомячки), то при вступлении их в митоз хромосомы могут объединиться в одну метафазную пластинку. После разделения такой клетки получится истинно гибридная клетка. Другие приемы позволяют конструировать клетки из разных по происхождению ядер и цитоплазмы. Так, разрушив актиновый компонент цитоскелета и подвергнув клетки центрифугированию, их можно разделить на две части: ядро с узким ободком цитоплазмы – кариопласт и на оставшуюся часть цитоплазмы – цитопласт. Затем, используя разные кариопласты и цитопласты, можно создавать разные комбинации реконструированных клеток.
Методы клеточной инженерии широко применяются не только в экспериментальной биологии, но и в биотехнологических целях. Например, при получении моноклональных антител используются клеточные гибриды между лимфоцитами иммунизированных животных и интенсивно размножающимися клетками миеломы. Полученные первичные дикарионы образуют истинные гибридные клетки, которые интенсивно размножаются за счет генома опухолевых миеломных клеток, и одновременно выделяют большое количество антител за счет работы генома иммунизированных лимфоцитов. Этот прием позволяет получать большое число гибридомных клеток, вырабатывающих большие количества необходимых антител.
РЕКОМЕНДАЦИИ ДЛЯ САМОПОДГОТОВКИ
Вопросы для обсуждения и размышления
Почему Ф. Энгельс в работе «Диалектика природы» назвал клеточную теорию одним из трех величайших открытий XIX века, написав: «Только со времени этого открытия стало на твердую почву исследование органических, живых продуктов природы – как сравнительная анатомия и физиология, так и эмбриология».
Какой смысл вкладывал Фридрих Энгельс в слова: «Покров тайны, окутавший процесс возникновения и роста и структуры организмов, был сорван. Непостижимое до этого времени чудо предстало в виде процесса, происходящего тождественному по существу для всех многоклеточных организмов закону».
Известный постулат Р.Вирхова гласит: «Каждая клетка происходит из клетки». Однако как в конце XIX, так и в XX веке некоторые ученые (М.Д.Лавровский, Н.П. Тишуткин, С.А. Усов и О.Б. Лепешинская) утверждали, что помимо клеток в составе организма существует живое вещество, способное к клеткообразованию. Можно ли согласиться с этой точкой зрения? Аргументируйте свою позицию.
Выпишите современную трактовку положений клеточной теории, постарайтесь к ним подобрать свои доказательства и примеры
В
 спомните название и назначение отдельных частей светового микроскопа, сделайте подписи к рисунку.
спомните название и назначение отдельных частей светового микроскопа, сделайте подписи к рисунку. Запомните правила микроскопии:
Смотрите в микроскоп одним глазом, другой глаз не закрывайте.
Одна рука на микрометрическом винте постоянно регулирует положение тубуса, что дает возможность рассмотреть всю толщу среза.
Другой рукой устанавливается и передвигается препарат на столике микроскопа.
С
Рис 15. Световой микроскоп
лева от микроскопа лежит тетрадь /альбом/ для зарисовки микропрепаратов.После окончания изучения препарата при большом увеличении, необходимо сменить большое увеличение на малое и только после этого снимать микропрепарат с предметного столика.
Оставлять микроскоп нужно всегда с подготовленным к работе объективом малого увеличения.
Помните! Оптика микроскопа боится пыли, поэтому микроскоп хранится в зачехленном состоянии.
Темы учебно-исследовательских работ
Кто изобрел микроскоп?
Значение работ Рудольфа Вирхова для дальнейшего развития биологии и медицины.
Современные методы изучения морфофизиологии клетки (на выбор).
Дополнительная литература
Атлас сканирующей электронной микроскопии клеток, тканей и органов. / Под. Ред. Волоковой, В.А. Шахламова, А.А. Миронова – М.: Медицина, 1987
О.В. Быков В.Л. Цитология и общая гистология (функциональная морфология клеток и тканей человека). – СПб.: СОТИС, 1998.
. Леви А., Сикевиц Ф. Структура и функции клетки. – М.: Мир, 1971.
Меркулов Г.А. Курс патологогистологической техники. – Изд-во Медицина, Ленинградское отделение, 1969.
Ченцов Ю.С. Общая цитология. – М.: Изд-во МГУ, 1995
Ченцов Ю.С. Введение в клеточную биологию. – М.: ИКЦ «Академкнига», 2004
Щелкунов С.И. Клеточная теория и учение о тканях. – Медгиз, Ленинградское отделение, 1958.
Ответы к вводному тесту.
Обмен веществ, рост, размножение.
Образована целлюлозой
Кислород.
Хромосомы.
Перенесения неблагоприятных условий внешней среды.
хитин
4
1
2
4
3
2,4,5
25
Просмотр содержимого документа
«клетка модуль 2»
Раздел 2. Общие принципы структурной организации клетки
При изучении этого раздела Вы узнаете:
общий план строения клетки, как основы единства органического мира, доказательства родства живой природы;
особенности строения и физиологии прокариотической клетки;
строение эукариотической клетки на микроскопическом и ультрамикроскопическом уровне;
морфофункциональные особенности клеточных органелл;
отличительные структурные особенности растительной и животной клетки.
При изучении этого раздела Вы научитесь:
на таблицах, схемах, микропрепаратах отличать растительные и животные клетки;
на рисунках, схемах, микропрепаратах и микрофотографиях отличать структурные компоненты клеток и описывать их.
ИНФОРМАЦИОННЫЙ БЛОК
Элементарной единицей всего живого является клетка - наименьшая жизнеспособная единица. По своему химическому составу все живые существа очень сходны. Основные компоненты всякой клетки - это нуклеиновые кислоты (ДНК, РНК), белки, жиры, углеводы. Изучение ультрамикроскопического строения различных типов клеток позволило выявить заметные различия между бактериями, цианобактериями с одной стороны и растениями и животными – с другой стороны. Различия между ними настолько велики, что эти две группы организмов отнесены к различным наиболее крупным таксономическим единицам – империи Прокариоты и Эукариоты.
Прокариот мы вправе рассматривать как реликтовые формы, сохранившиеся (и процветающие!) с самых ранних времен биологической эволюции. А появление эукариот, возникших, более всего вероятно, из прокариот - как величайший скачок эволюции органического мира.
Для наиболее полного усвоения учебного материала об особенностях строения прокариотических клеток Вам необходимо вспомнить следующие темы из курса биологии (6 класс): основы классификации бактерий, особенности строения бактериальной клетки, значение бактерий в природе и жизни человека.
В настоящее время бактерии выделены в отдельное царство. Наиболее распространённой системой классификации бактерий является система Берги.
Бактерии - одноклеточные организмы, лишенные хлорофилла и размножающиеся, в основном простым делением, поэтому их раньше относили к дробянкам. По форме различают шаровидные бактерии, палочковидные и извитые. Шаровидные бактерии называются кокки (coccus - зерно). Размеры кокков 1-1,5 мкм. Кокки различаются между собой в зависимости от расположения клеток в мазке (это определяется особенностями их деления). Одиночные кокки н азываются микрококки, в основном это сапрофитные обитатели воздуха, воды и почвы, среди них мало болезнетворных организмов. Диплококки лежат в мазке попарно, среди них есть возбудители опасных для человека инфекционных заболеваний: пневмококки, гонококки и менингококки. Стрептококки в мазке расположены в виде цепочки, это возбудители гнойных инфекций – ангины (тонзиллита), пневмонии, отита, послеродового сепсиса и др. Тетракокки в мазке лежат по четыре клетки, и как правило это не патогенные (не вызывающие заболевания) микробы. Стафилококки в мазке расположены в виде беспорядочных скоплений, напоминающих гроздья винограда. Стафилококки, так же как и стрептококки, являются пиогенными (гноеродными) микроорганизмами. Они являются возбудителями отита, гайморита, остеомиелита, мастита, сепсиса и многих других гнойных заболеваний.
азываются микрококки, в основном это сапрофитные обитатели воздуха, воды и почвы, среди них мало болезнетворных организмов. Диплококки лежат в мазке попарно, среди них есть возбудители опасных для человека инфекционных заболеваний: пневмококки, гонококки и менингококки. Стрептококки в мазке расположены в виде цепочки, это возбудители гнойных инфекций – ангины (тонзиллита), пневмонии, отита, послеродового сепсиса и др. Тетракокки в мазке лежат по четыре клетки, и как правило это не патогенные (не вызывающие заболевания) микробы. Стафилококки в мазке расположены в виде беспорядочных скоплений, напоминающих гроздья винограда. Стафилококки, так же как и стрептококки, являются пиогенными (гноеродными) микроорганизмами. Они являются возбудителями отита, гайморита, остеомиелита, мастита, сепсиса и многих других гнойных заболеваний.
Палочковидные бактерии (от гр. bacteriae - палочка) имеют цилиндрическую форму и в мазке располагаются обычно одиночно, но иногда попарно (диплобактерии) или в виде цепочек (стрептобактерии). Палочки, образующие споры, называются бациллами. Спорообразующие анаэробы, живущие в безкислородной срезе, называются клостридиями. Среди бактерий много возбудителей инфекционных заболеваний: чумы, сибирской язвы, столбняка, гангрены, дифтерии, кишечной инфекции.
И звитые формы бактерий имеют вид спирали, с завитками - это спириллы. Спириллы - безвредные микроорганизмы, живущие в сточных или загрязненных водах, а также в гниющих отбросах. Только Spirillum minus вызывает у человека болезнь укуса крыс - содоку.
звитые формы бактерий имеют вид спирали, с завитками - это спириллы. Спириллы - безвредные микроорганизмы, живущие в сточных или загрязненных водах, а также в гниющих отбросах. Только Spirillum minus вызывает у человека болезнь укуса крыс - содоку.
В аааа б в
Спирохеты - бактерии, которые строением клеток и способом передвижения отличаются от всех других бактерий. В клетках спирохет можно различать три главных компонента: спирально извитой протоплазматический цилиндр, осевые фибриллы и наружную оболочку.
Бактериальная клетка состоит из клеточной оболочки, цитоплазматической мембраны и цитоплазмы, которая содержит ядерное вещество и различные включения и органеллы.
. Особенности строения и физиологии прокариотической клетки
Строение и некоторые функциональные особенности прокариотичсских клеток мы рассмотрим более подробно. Прежде чем перейти к деталям, отметим основные признаки, отличающие прокариотическую клетку от эукариотической. Клетки прокариот очень малы. Подавляющее большинство бактерий имеет размеры не более 1 мкм толщиной и 5 мкм длиной.
ДНК прокариотической клетки не окружена ядерной мембраной, а органеллы типа митохондрий и хлоропластов отсутствуют. Область ядерного вещества, видимая на электронной микрофотографии ультратонкого среза клетки в виде сетчатой структуры из тонких нитей, граничит с заполненной рибосомами цитоплазмой. У многих бактерий впячивания плазматической мембраны образуют структуры во внутреннем пространстве цитоплазмы - это мезосомы, участвующие в реакциях энергетического обмена и процессе деления.
В цитоплазме бактерии расположены рибосомы - гранулы, состоящие из р-РНК и белка. Рибосомы осуществляют синтез белка. Рибосомы прокариот меньше, чем цитоплазматические рибосомы эукариот, они относятся к типу 70S. Скопления рибосом называются полисомы.
В
ся генетическая информация клетки прокариот содержится в одной единственной кольцевой нити ДНК - нуклеоиде - «бактериальной хромосоме». Нуклеоид содержит гистоноподобные белки. Молекула ДНК у всех до сих пор исследованных бактерий имеет вид кольцевой нити длиной 0,25-3 мм. У многих бактерий были обнаружены внехромосомные, замкнутые в кольцо молекулы ДНК - плазмиды. Информация, содержащаяся в плазмидах, не является необходимой для клеток и чаще всего несет гены устойчивости бактерии к различным факторам, а также половой F-фактор (фактор фертильности). Бактерии, как правило, размножаются путем бинарного деления клетки. Делению клетки предшествует удвоение или репликация бактериальной ДНК.
Клеточная оболочка прокариот двух или трехслойна. Капсула – это наружный, слизистый слой (химический состав капсулы: 98 % воды, остальное приходится на долю полисахаридов), у различных бактерий капсула имеет разное развитие. Одни бактерии образуют капсулу, когда попадают в организм хозяина (например, пневмококки), другие обладают постоянной капсулой.
К
апсула предохраняет бактерию от действия фагоцитов и других иммунных факторов организма хозяина. Клеточная стенка - оболочка, которая оделяет микробную клетку от окружающей среды. Клеточная стенка прочная и эластичная, имеет толщину 10-20 нм. Прочность стенке придает сеть микрофибрилл из пептидогликана (муреина), погруженных в матрикс. В матриксе обнаружены полисахариды, липиды, белки. Обладая избирательной проницаемостью, клеточная стенка обеспечивает прохождение в клетку различных веществ: воды, глюкозы, аминокислот, жирных кислот и др. Более крупные молекулы органических веществ в клетку проникнуть не могут без предварительного расщепления ферментами, выделяемыми клеткой. Цитоплазматическая мембрана (самый внутренний слой) имеет типичное строение элементарной мембраны. При плазмолизе тело клетки отделяется от клеточной стенки. Клеточная стенка под действием лизоцима или пенициллина разрушается. Бактерии, целиком лишенные клеточной стенки, называются протопластами, а имеющие остатки ее - сферопластами. Эти образования, покрытые тонкой цитоплазматической мембраной, могут существовать только в специальных осмотически уравновешенных средах (5-20 % сахарозы).
Многие бактерии содержат включения - гранулы крахмала, белка, гликогена, зерна волютина (что часто является важным диагностическим признаком).
Н екоторые бактерии, попадая в неблагоприятные условия, образуют споры внутри тела. Спора устойчива к высушиванию, действию низких температур, химических и дезинфицирующих веществ. Спорообразование свойственно бациллам. Спорообразование - это
екоторые бактерии, попадая в неблагоприятные условия, образуют споры внутри тела. Спора устойчива к высушиванию, действию низких температур, химических и дезинфицирующих веществ. Спорообразование свойственно бациллам. Спорообразование - это
процесс обособления ядерного вещества и части цитоплазмы оболочкой, состоящей из липидов солей кальция и минимальным содержанием воды. Спорообразование длится 18-20 часов.
Оставшаяся (вегетативная) часть клетки вскоре отмирает. В споре затухают все обменные
процессы. У различных видов бактерий споры различаются по размеру, форме и по расположению. Возбудитель столбняка, например, образует круглую спору на самом конце палочки. У возбудителя сибирской язвы спора расположена посередине. Возбудитель ботулизма имеет очень большую спору на конце клетки.
М ногие прокариоты подвижны - они перемещаются путем плавания или скольжения. Органами движения плавающих бактерий служат особые «бактериальные» жгутики. Эти жгутики устроены значительно проще, чем у эукариот, и состоят из одной-единственной фибриллы.
ногие прокариоты подвижны - они перемещаются путем плавания или скольжения. Органами движения плавающих бактерий служат особые «бактериальные» жгутики. Эти жгутики устроены значительно проще, чем у эукариот, и состоят из одной-единственной фибриллы.
Физиологическими особенностями прокариотической клетки являются: аэробный и анаэробный тип дыхания, синтез белка (транскрипция и трансляция) в одних компартментах клетки.
Табл. 1 Сравнительная характеристика прокариотических и эукариотисческих организмов
| Характеристика | Прокариоты | Эукариоты |
| Организмы | Бактерии, цианобактерии, архибактерии. | Грибы, растения, животные. |
| Клеточная организация | Одноклеточные | Одноклеточные и многоклеточные с клеточной дифференцировкой |
| Клеточные стенки | Плотная, достаточно жесткая, содержит полисахариды, аминокислоты и липиды. Основной компонент – муреин. | У зеленых растений и грибов клеточная стенка жесткая содержит полисахариды. Основной компонент клеточной стенки растений - целлюлоза, у грибов - хитин. |
| ДНК | Кольцевая ДНК в цитоплазме. Нет ядра, ограниченного мембраной, ДНК с белковым комплексом образует нуклеоид. | Длинные ДНК с большим количеством некодирующих участков организованы в хромосомы,окружены ядерной оболочкой. Есть ядрышки. |
| Цитоплазма | Отсутствие цитоскелета, движения цитоплазмы, эндо- и экзоцитоза | Имеется микротробекулярный цитоскелет, активное движение цитоплазмы, эндоцитоз и экзоцитоз. |
| Органеллы | Органелл мало. Присутствуют мезосомы и рибосомы | Имеются мембранные органеллы - эндоплазматическая сеть, митохондрии, пластиды, комплекс Гольджи, лизосомы, пероксисомы; и немебранные органеллы - рибосомы, центриоли. |
| РНК и белки | РНК и белки синтезируются в одном компартменте - цитоплазме. | Синтез и созревание (процессинг) РНК происходит в ядре, синтез белков в цитоплазме. |
| Жгутики | Простые, состоят из одной или нескольких фибрилл. | Сложные, с расположением микротрубочек типа 9+2. |
| Деление | Бинарное деление. | Митоз (или мейоз), амитоз. |
| Метаболизм | Анаэробный или аэробный | Аэробный или вторично анаэробный |
29
Просмотр содержимого документа
«клетка модуль 3»
2.2. Общий план строения эукариотической клетки
Клетки многоклеточного животного организма разнообразны. Значительно различаются по форме, размерам, по строению и функциям клетки крови, мышечные и нервные клетки, сперматозоиды и клетки желез.
Клетки растений также достаточно разнообразны: это клетки эпидермы и мезофила, ситовидные трубочки и волокна колленхимы.
О
днако исходя из постулата Р. Вирхова о гомологичности клеток, можно говорить о едином плане строения всех клеток: они состоят из поверхностного аппарата, ядра и цитоплазмы.
эукариотическая клетка

| поверхностный аппарат клетки
| Внутреннее содержимое клетки (протопласт) | |||||
| Цитоплазматическая мембрана | Ядро
| Цитоплазма | ||||
| Ядерная оболочка | Нуклеоплазма | Гиалоплазма | Органеллы | Включения | ||
| Кортикальный слой цитоплазмы
Гликокаликс (животные клетки) или
Клеточная стенка (растительные клетки) |
Хроматин Ядерный матрикс Ядрышко
|
Мембраные |
Немембранные |
Трофические Секреторные Экскреторные Пигментные | ||
| ЭПС – гладкая и гранулярная Лизосомы Митохондрии Пластиды Комплекс Гольджи Глиоксисомы Синаптические пузырьки | Рибосомы Центриоли
| |||||
Все эукариотические клетки отличаются значительным развитием мембранных структур.
2.2.1. Морфофункциональные особенности клеточных мембран
Клеточные мембраны (6-10 нм) - важнейший компонент живого содержимого клетки, выполняют барьерную, транспортную и рецепторную функции.
Все без исключения клеточные мембраны построены по общему принципу: это тонкие липопротеидные пленки, состоящие из двойного слоя липидных молекул, в который включены молекулы белка. На долю липидов (жироподобных веществ) приходится 20-80%, на долю белков 20-80%. В состав многих мембран входят углеводы, количество которых достигает 2-10%. К липидам относится большая группа органических веществ, обладающих плохой растворимостью в воде (гидрофобность) и хорошей растворимостью в органических растворителях (липофильность). Характерными представителями липидов, встречающихся в клеточных мембранах, являются фосфолипиды (глицерофосфатиды), гликолипиды, сфингомиелины и холестерин. В растительных клетках холестерин не обнаружен, его там заменяют фитостерины (эргостерол). Характерной особенностью липидов мембран является разделение их молекулы на неполярные хвосты, состоящие из жирных кислот, и заряженные полярные головки, образованные многоатомными спиртами. На поверхности воды растворы полярных липидов, растекаясь, образуют мономолекулярную пленку, в которой в водную фазу будут направлены заряженные (гидрофильные) головки, а неполярные хвосты будут обращены в сравнительно гидрофобную воздушную фазу. Смеси разных липидов формируют бимолекулярные слои или мембраны толщиной около 3,5 нм, где периферические зоны слоя, смотрящие в воду, будут содержать полярные головки, а незаряженные хвосты будут образовывать общую гидрофобную центральную часть образовавшейся мембраны. Таким образом, способность липидов самопроизвольно образовывать мембранные структуры определяется их химической структурой. Как искусственные, так и естественные мембраны не представляют собой плоские слои, они всегда замкнуты сами на себя и образуют полые вакуоли, пузырьки, плоские замкнутые мешки или трубчатые образования. Липидные мембраны служат непроницаемым барьером для любых заряженных молекул, даже для ионов солей. Это определяет основное функциональное свойство мембран - служить преградой для свободной диффузии через слой липидов. Это свойство может быть использовано для практических целей. Так при смешивании липидов в водной среде образуется масса полых мембранных пузырьков - липосом. Жидкость, попавшая внутрь этих пузырьков, уже не может свободно обмениваться с жидкостью, находящейся снаружи. Таким образом, искусственные мембраны липосом можно «загрузить» лекарственными веществами, которые могут в нужных концентрациях поступать к клеткам.
Белки – второй компонент клеточных мембран. Количество белков в разных мембранах может быть различным. Так в мембранах митохондрий доля белков составляет около 75%, а в плазматической мембране клеток миелиновой оболочки - около 25%. Белковые молекулы как бы вкраплены в билипидный слой мембраны. Часть из них связана с липидными головками с помощью ионных связей и поэтому легко экстрагируется из мембран растворами солей. Легко экстрагируемые белки расположены на поверхности мембран и их называют периферийными. Большая часть белков взаимодействует с липидами в составе мембран на основе гидрофобных связей. Оказалось, что многие мембранные белки состоят как бы из двух частей: из участков, богатых полярными аминокислотами, и участков, обогащенных неполярными аминокислотами. Такие белки в липидных слоях мембран располагаются так, что их неполярные участки погружены в «жирную» часть мембраны, где находятся гидрофобные участки липидов. Полярная часть таких белков взаимодействует с головками липидов и обращена в сторону водной фазы, поэтому такие белки, связанные с липидами путем гидрофобных взаимодействий, практически не экстрагируются в водных фазах. Эти белки мембран называют интегральными. Размер интегральных мембранных белков в среднем равен 8 нм, но встречаются крупные белки - до 35 нм величиной (белок тилакоидов хлоропластов). Интересно, что большая часть липидных молекул (70%) не связана с белками, так что белковые молекулы как бы плавают в «липидном озере». Липидные молекулы также двигаются вдоль липидного слоя, могут вращаться вокруг своей оси. Гарт Л. Николс и С. Дж. Синджер в 1972 г. предложили жидкостно-мозаичную модель строения элементарной мембраны.
Н
есмотря на поразительную схожесть строения различных мембран, построенных по принципу липидного бислоя с вмонтированными в него белками, физические и химические свойства разных мембран различны. Это связано с тем, что в разных мембранах общий состав липидов значительно различается, что определяет особые свойства мембран.
Разные мембраны клетки могут отличаться друг от друга по количеству липидов. Так, плазматическая мембрана содержит 35-40% липидов, а мембраны митохондрий - 27-29%. Самое высокое содержание липидов в плазматической мембране шванновских клеток, образующих миелиновую оболочку нервов, - до 80%. Было обнаружено, что клеточные мембраны сильно отличаются друг от друга по составу липидов. Так, плазматические мембраны клеток животных богаты холестерином (до 30%), в то время как мембраны митохондрий, наоборот, богаты фосфолипидами и бедны холестерином.
Особенно отличаются мембраны по составу белков, которые, главным образом, определяют функциональные свойства мембран. По биологической роли мембранные белки можно разделить на группы: ферменты, рецепторные белки, структурные и транспортные белки.
Набор ферментов в составе мембран может быть очень велик и разнообразен (например, в плазматической мембране клеток печени обнаружено не менее 24 различных ферментов). В разных мембранах существует характерный набор ферментов. Например, в плазматической мембране, как и во многих других, локализуется K+-Na+-зависимая АТФаза, участвующая в транспорте ионов. В митохондриях специфическим является набор белков - переносчиков электронов и фермент АТФ-синтетаза, обеспечивающие окислительное фосфорилирование и синтез АТФ.
Рецепторные белки специфически связываются с теми или иными веществами и как бы их «узнают». Это белки-рецепторы для гормонов, для узнавания поверхности соседних клеток, вирусов, фагов у бактерий и т.д. К этой группе относятся фоторецепторные белки. Рецепторные белки входят в состав любых мембран.
Со стороны цитоплазмы мембраны связаны через примембранные или собственно мембранные интегральные белки с разнообразными белковыми структурами цитоплазмы. К ним относятся в первую очередь компоненты цитоскелета. Это позволяет не только сделать мембраны более жесткими, но и обеспечивает подвижность мембран, создавая возможности для их транспортных функций. Во многих эпителиальных клетках специальные белки плазматической мембраны связываются с элементами цитоскелета и участвуют в образовании целого ряда межклеточных соединений (десмосомы, адгезивный контакт и др.).
2.2.2. Поверхностный аппарат клетки
Плазмолемма среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, ограничивающая клетку снаружи. Плазматическая мембрана исполняет роль барьера между внутриклеточным содержимым и внешней средой; она ограничивает свободный поток низко- и высокомолекулярных веществ в обе стороны через мембрану. Плазмолемма - это структура «узнающая» различные химические вещества и регулирующая избирательно транспорт этих веществ в клетку и из нее. Плазмолемма участвует в делении клетки, в образовании ресничек и микроворсинок, а так же в формировании межклеточных контактов.
Г ликокаликс представляет собой внешний по отношению к липопротеидной мембране слой, содержащий полисахаридные цепочки мембранных интегральных белков - гликопротеидов. Эти цепочки содержат такие углеводы как манноза, глюкоза и др. Углеводные гетерополимеры образуют ветвящиеся цепочки, между которыми могут располагаться выделенные из клетки гликолипиды и протеогликаны. Слой гликокаликса сильно обводнен, имеет желеподобную консистенцию, что значительно снижает в этой зоне скорость диффузии различных веществ. Здесь же могут «застревать» выделенные клеткой гидролитические ферменты, участвующие во внеклеточном расщеплении полимеров (внеклеточное пищеварение) до мономерных молекул, которые затем транспортируются в цитоплазму через плазматическую мембрану.
ликокаликс представляет собой внешний по отношению к липопротеидной мембране слой, содержащий полисахаридные цепочки мембранных интегральных белков - гликопротеидов. Эти цепочки содержат такие углеводы как манноза, глюкоза и др. Углеводные гетерополимеры образуют ветвящиеся цепочки, между которыми могут располагаться выделенные из клетки гликолипиды и протеогликаны. Слой гликокаликса сильно обводнен, имеет желеподобную консистенцию, что значительно снижает в этой зоне скорость диффузии различных веществ. Здесь же могут «застревать» выделенные клеткой гидролитические ферменты, участвующие во внеклеточном расщеплении полимеров (внеклеточное пищеварение) до мономерных молекул, которые затем транспортируются в цитоплазму через плазматическую мембрану.
В электронном микроскопе гликокаликс имеет вид рыхлого волокнистого слоя, толщиной 3-4 нм, покрывающего всю поверхность клетки. Он обнаружен практически у всех животных клеток, но степень его выраженности различна. Особенно хорошо гликокаликс развит в щеточной каемке клеток всасывающего эпителия кишечника.
Механическая устойчивость плазматической мембраны, кроме того, обеспечивается структурой примыкающего к ней со стороны цитоплазмы кортикального слоя и внутриклеточных фибриллярных структур. Кортикальный (от слова cortex - кора, кожица) слой цитоплазмы, лежащий в тесном контакте с плазмолеммой, имеет ряд особенностей. Здесь, при толщине его в 0,1-0,5 мкм, отсутствуют рибосомы и мембранные пузырьки, но в большом количестве встречаются фибриллярные элементы цитоплазмы - микрофиламенты и, часто, микротрубочки. Основным фибриллярным компонентом кортикального слоя является сеть микрофибрилл актиновых белков и вспомогательных белков, необходимых для движения участков цитоплазмы.
Транспорт ионов через плазмалемму проходит за счет участия в этом процессе мембранных транспортных белков - пермеаз. Эти белки могут вести транспорт в одном направлении одного вещества (унипорт), или нескольких веществ одновременно (симпорт), или же вместе с импортом одного вещества выводить из клетки другое (антипорт). Так, например, глюкоза может входить в клетки симпортно вместе с ионом Na+.
Т ранспорт ионов может происходить по градиенту концентрации – пассивно, без дополнительной затраты энергии. Так, например, в клетку проникает ион Na+ из внешней среды, где его концентрация выше, чем в цитоплазме. В случае пассивного транспорта некоторые мембранные транспортные белки образуют молекулярные комплексы, каналы, через которые растворенные молекулы проходят через мембрану за счет простой диффузии по градиенту концентрации. Часть этих каналов открыта постоянно, а другая часть может закрываться или открываться в ответ либо на связывание с сигнальными молекулами, либо на изменение внутриклеточной концентрации ионов. В других случаях специальные мембранные белки-переносчики избирательно связываются с тем или иным ионом и переносят его через мембрану - это облегченная диффузия.
ранспорт ионов может происходить по градиенту концентрации – пассивно, без дополнительной затраты энергии. Так, например, в клетку проникает ион Na+ из внешней среды, где его концентрация выше, чем в цитоплазме. В случае пассивного транспорта некоторые мембранные транспортные белки образуют молекулярные комплексы, каналы, через которые растворенные молекулы проходят через мембрану за счет простой диффузии по градиенту концентрации. Часть этих каналов открыта постоянно, а другая часть может закрываться или открываться в ответ либо на связывание с сигнальными молекулами, либо на изменение внутриклеточной концентрации ионов. В других случаях специальные мембранные белки-переносчики избирательно связываются с тем или иным ионом и переносят его через мембрану - это облегченная диффузия.
В клетках существуют мембранные белковые переносчики, которые работают против градиента концентрации, затрачивая при этом энергию АТФ. Такой тип работы носит название активного транспорта, и он осуществляется с помощью белковых ионных насосов. В плазматической мембране находится двухсубъединичная молекула (K+ + Na+)-насоса, которая одновременно является и АТФазой. Этот насос при работе откачивает за один цикл 3 иона Na+ и закачивает в клетку 2 иона K+ против градиента концентрации. При этом затрачивается одна молекула АТФ, идущая на фосфорилирование АТФазы, в результате чего Na+ переносится через мембрану из клетки, а K+ получает возможность связаться с белковой молекулой и затем переносится в клетку.
клетках существуют мембранные белковые переносчики, которые работают против градиента концентрации, затрачивая при этом энергию АТФ. Такой тип работы носит название активного транспорта, и он осуществляется с помощью белковых ионных насосов. В плазматической мембране находится двухсубъединичная молекула (K+ + Na+)-насоса, которая одновременно является и АТФазой. Этот насос при работе откачивает за один цикл 3 иона Na+ и закачивает в клетку 2 иона K+ против градиента концентрации. При этом затрачивается одна молекула АТФ, идущая на фосфорилирование АТФазы, в результате чего Na+ переносится через мембрану из клетки, а K+ получает возможность связаться с белковой молекулой и затем переносится в клетку.
Такая постоянная работа пермеаз и насосов создает в клетке постоянную концентрацию ионов и низкомолекулярных веществ. В сочетании с активным транспортом ионов через плазматическую мембрану происходит транспорт различных сахаров, нуклеотидов и аминокислот.
Макромолекулы, такие как белки, нуклеиновые кислоты, полисахариды, липопротеидные комплексы и другие, сквозь клеточные мембраны не проходят. В клетку же или из одного мембранного компартмента в другой макромолекулы попадают заключенными внутри вакуолей или везикул. Такой везикулярный перенос можно разделить на два вида: экзоцитоз - вынос из клетки макромолекулярных продуктов и эндоцитоз - поглощение клеткой макромолекул.
При эндоцитозе определенный участок плазмалеммы захватывает, как бы обволакивает, внеклеточный материал, заключает его в мембранную вакуоль, возникшую за счет впячивания плазматической мембраны. В такую первичную вакуоль могут попадать любые биополимеры, части клеток или даже целые клетки, где затем и распадаются до мономеров. Основное биологическое значение эндоцитоза - это получение строительных блоков за счет внутриклеточного переваривания, которое осуществляется после слияния первичной эндосомы с лизосомой.
Эндоцитоз формально разделяют на пиноцитоз и фагоцитоз. Фагоцитоз - захват и поглощение клеткой крупных частиц (иногда даже клеток или их частей) - был впервые описан И.И. Мечниковым. Фагоцитоз, способность захватывать клеткой крупные частицы, встречается среди клеток животных как одноклеточных (например, амебы, некоторые хищные инфузории), так и для специализированных клеток многоклеточных животных - фагоцитов. Пиноцитоз вначале определялся как поглощение клеткой воды или водных растворов разных веществ. Сейчас известно, что как фагоцитоз так и пиноцитоз протекают очень сходно, и поэтому употребление этих терминов может отражать лишь различия в объемах, массе поглощенных веществ. Общее для этих процессов то, что поглощенные вещества на поверхности плазматической мембраны окружаются мембраной в виде вакуоли - эндосомы, которая перемещается внутрь клетки.
Дальнейшая судьба эндосом может быть различной: часть из них может возвращаться к поверхности клетки и сливаться с ней, но большая часть вступает в процесс внутриклеточного пищеварения. Первичные эндосомы содержат в основном захваченные в жидкой среде чужеродные молекулы и не содержат гидролитических ферментов. Эндосомы могут сливаться друг с другом, при этом увеличиваясь в размере. Они затем сливаются с первичными лизосомами, которые вводят в полость эндосом ферменты, расщепляющие (гидролизующие) различные биополимеры.
Фагоцитоз является вариантом эндоцитоза и связан с поглощением клеткой крупных агрегатов макромолекул вплоть до живых или мертвых клеток. При фагоцитозе происходит образование больших эндоцитозных вакуолей - фагосом, которые затем, сливаясь с лизосомами, образуют фаголизосомы.
Плазматическая мембрана принимает участие в выведении веществ из клетки с помощью экзоцитоза - процесса, обратного эндоцитозу. В случае экзоцитоза, внутриклеточные продукты, заключенные в вакуоли или пузырьки и отграниченные от гиалоплазмы мембраной, подходят к плазматической мембране. В местах их контактов плазматическая мембрана и мембрана вакуоли сливаются, и пузырек опустошается в окружающую среду. С экзоцитозом связано выделение синтезированных в клетке разнообразных веществ. Клетки могут вырабатывать и выделять во внешнюю среду низкомолекулярные соединения (ацетилхолин, биогенные амины и др.), а также макромолекулы: пептиды, белки, липопротеиды, пептидогликаны и др.. Выделенные из клеток путем экзоцитоза гидролитические ферменты могут сорбироваться в слое гликокаликса и обеспечивать примембранное внеклеточное расщепление различных биополимеров и органических молекул. Огромное значение примембранное неклеточное пищеварение имеет для животных. Было обнаружено, что в кишечном эпителии млекопитающих в зоне щеточной каемки всасывающего эпителия, особенно богатой гликокаликсом, обнаруживается огромное количество разнообразных ферментов.
2.2.3. Цитоплазма
Цитоплазма - это тот компонент клетки, который остается, если исключить ядро. Цитоплазма клетки многокомпонентна. Даже в световой микроскоп в цитоплазме живой клетки видны вкрапления, неоднородности, частички, но особенно неоднородность цитоплазмы видна при изучении ее в электронном микроскопе.
Формально структуру цитоплазмы подразделяют на три части: органеллы, включения, гиалоплазма (основная плазма или цитозоль).
О рганеллы - обязательные для любой клетки компоненты, без которых клетка просто не может поддерживать свое существование, они имеют постоянную структуру и выполняют определенные функции. Включения – это необязательные компоненты, которые представляют собой или отложения запасных веществ (гликоген, желточные гранулы) или скопление продуктов метаболизма (пигменты, кристаллы солей и др. в растительных клетках).
рганеллы - обязательные для любой клетки компоненты, без которых клетка просто не может поддерживать свое существование, они имеют постоянную структуру и выполняют определенные функции. Включения – это необязательные компоненты, которые представляют собой или отложения запасных веществ (гликоген, желточные гранулы) или скопление продуктов метаболизма (пигменты, кристаллы солей и др. в растительных клетках).
Органеллы и включения погружены в гиалоплазму - жидкую фазу цитоплазмы клетки.
2.2.3.1. ГиалоплазмаТермины гиалоплазма (от hyaline - прозрачный), основная плазма, матрикс цитоплазмы или цитозоль, обозначают очень важную часть клетки, ее истинную внутреннюю среду. Гиалоплазма - не просто разбавленный водный раствор органических соединений: его состав весьма сложен, а консистенция приближается к гелю (желе). Гели - это структурированные коллоидные системы с жидкой дисперсной средой. Частицы дисперсной фазы соединены между собой в рыхлую пространственную сетку, которая содержит в своих ячейках дисперсную среду, лишая текучести систему в целом. Гель гиалоплазмы под воздействием внешних условий (температура, давление) или внутренних факторов может переходить в менее вязкую, более жидкую фазу - в золь или раствор. Такие гель-золь переходы очень характерны для гиалоплазмы. Отдельные зоны гиалоплазмы могут менять свое агрегатное состояние в зависимости от условий или функции. Отдельные молекулы белков-тубулинов могут быть диспергированы в гиалоплазме, но в определенные моменты они начинают собираться и строить длинные трубчатые структуры - микротрубочки. Этот процесс самосборки микротрубочек обратим: при изменении условий последние распадаются до мономерных молекул тубулинов. Таким же образом в гиалоплазме могут возникать и распадаться различные фибриллярные, нитчатые комплексы белковых молекул.
Функциональное значение гиалоплазмы очень велико. Здесь находятся ферменты, участвующие в синтезе аминокислот, нуклеотидов, жирных кислот, сахаров. В гиалоплазме происходит синтез и отложение запасного полисахарида гликогена, накопление запасных жировых капель. Здесь же происходят процессы гликолиза и синтез части АТФ.
В гиалоплазме на рибосомах и полирибосомах, не связанных с мембранами ЭПС, происходит синтез белков, которые транспортируются в различные участки клетки. Здесь осуществляется синтез всех белков клеточного ядра, большей частью белков митохондрий и пластид. Кроме структурных белков и ферментов в цитозоле в растворенном состоянии содержится огромное количество аминокислот, нуклеотидов и других строительных блоков биополимеров, а также множество метаболитов - промежуточных продуктов, возникающих при синтезе и распаде макромолекул. Гиалоплазма содержит большое количество ионов неорганических соединений, таких как Na+, K+, Ca2+, Cl-, HCO3-, HPO42- и др. При этом концентрация этих ионов строго регулируется мембранными компонентами клетки.
Формально, по морфологическим признакам, органеллы можно разделить на две группы: мембранные и немембранные. Мембранные органеллы так же представлены двумя вариантами: одномембранные и двумембранные. К первым относятся органеллы вакуолярной системы – эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы и другие специализированные вакуоли. К двумембранным органеллам относятся митохондрии и пластиды. К немембранным органеллам принадлежат рибосомы, клеточный центр клеток низших растений и животных. Клеточный скелет или цитоскелет - постоянный компонент клетки, его выраженность может значительно меняться в течение клеточного цикла.
2.2.3.2. Эндоплазматическая сеть
Эндоплазматическая сеть – ЭПС (ретиклум ЭР) - сквозная сеть двухконтурных мембран, соединенных друг с другом, образующая каналы, вакуоли, мешочки и трубочки. Эта внутриклеточная мембранная структура была открыта в 1945 году К. Портером. В 50-х гг., при использовании метода ультратонких срезов удалось выяснить структуру эндоплазматического ретикулюма и обнаружить его неоднородность. Подобный электронно-микроскопический анализ позволил выделить два типа ЭПС: гранулярная (шероховатая) и агранулярная или гладкая ЭПС.
Г ранулярная ЭПС представлена замкнутыми мембранами, которые образуют на сечениях вытянутые мешки, цистерны или же имеют вид узких (около 20 нм) каналов. Отличительной особенностью гранулярной ЭПС, является то, что ее мембраны со стороны гиалоплазмы покрыты мелкими (около 20 нм) темными, почти округлыми частицами, гранулами - рибосомами. На мембранах рибосомы расположены в виде полисом (множество рибосом, объединенных одной информационной РНК). Это синтезирующие белок рибосомы прикрепляются к мембранам большой субъединицей. Гранулярная ЭПС может в клетках быть представлена или в виде редких разрозненных мембран или же в виде скоплений - это эргастоплазма. Первый тип гранулярноой ЭПС характерен для малодифференцированных клеток или клеток с низкой метаболической активностью. Эргастоплазма характерна для клеток, активно синтезирующих белки. В клетках печени каналы гранулярной ЭПС собраны в отдельные зоны - тельца Берга, а в нервных клетках формирует тигроид или вещество Ниссля. Количество рибосом на ЭПС четко связано с ее синтетической активностью. Например, на мембранах ЭПС в клетке несекретирующей молочной железы связывается до 25% клеточных рибосом, в лактирующей железе их количество возрастает до 70%. Уменьшение числа рибосом наблюдается и при различных патологических состояниях клеток (при алкогольном хроническом отравлении происходит уменьшение числа связанных рибосом на 25%). Рибосомы, связанные с мембранами ЭПС, участвуют в синтезе белков, выводимых из данной клетки, т.е. «экспортируемых» белков.
ранулярная ЭПС представлена замкнутыми мембранами, которые образуют на сечениях вытянутые мешки, цистерны или же имеют вид узких (около 20 нм) каналов. Отличительной особенностью гранулярной ЭПС, является то, что ее мембраны со стороны гиалоплазмы покрыты мелкими (около 20 нм) темными, почти округлыми частицами, гранулами - рибосомами. На мембранах рибосомы расположены в виде полисом (множество рибосом, объединенных одной информационной РНК). Это синтезирующие белок рибосомы прикрепляются к мембранам большой субъединицей. Гранулярная ЭПС может в клетках быть представлена или в виде редких разрозненных мембран или же в виде скоплений - это эргастоплазма. Первый тип гранулярноой ЭПС характерен для малодифференцированных клеток или клеток с низкой метаболической активностью. Эргастоплазма характерна для клеток, активно синтезирующих белки. В клетках печени каналы гранулярной ЭПС собраны в отдельные зоны - тельца Берга, а в нервных клетках формирует тигроид или вещество Ниссля. Количество рибосом на ЭПС четко связано с ее синтетической активностью. Например, на мембранах ЭПС в клетке несекретирующей молочной железы связывается до 25% клеточных рибосом, в лактирующей железе их количество возрастает до 70%. Уменьшение числа рибосом наблюдается и при различных патологических состояниях клеток (при алкогольном хроническом отравлении происходит уменьшение числа связанных рибосом на 25%). Рибосомы, связанные с мембранами ЭПС, участвуют в синтезе белков, выводимых из данной клетки, т.е. «экспортируемых» белков.
Роль гранулярной ЭПС заключается не только в участии в синтезе белков на рибосомах его мембран, но и в процессе обособления этих синтезированных белков, в их изоляции от других белков клетки. Эта функциональная особенность гранулярной ЭПС обеспечивает выделение синтезированных белков с помощью вакуолей комплекса Гольджи. В гранулярной ЭПС п роисходит синтез белков, которые, встраиваясь в ее мембрану, становятся интегральными мембранными белками.
роисходит синтез белков, которые, встраиваясь в ее мембрану, становятся интегральными мембранными белками.
Сегодня можно сказать, что важнейшей функцией гранулярной ЭПС, вне зависимости от специализации или тканевой принадлежности клеток, является функция образования, построения клеточных мембран, которая заключается в том, что элементы гранулярной ЭПС синтезируют все мембранные белки, синтезируют липидный компонент мембран, но, кроме того, именно здесь происходит сборка липопротеидных мембран. От того, какие интегральные и периферические белки будут синтезироваться на рибосомах ЭПС, и от того, какие фосфолипиды будут синтезироваться и включаться в мембрану, будет зависеть тип образующегося нового участка мембраны: будет ли он компонентом гладкой ЭПС, мембран комплекса Гольджи, лизосомы. или плазматической мембраны.
Участки гранулярной ЭПС, которые расположены в зоне около комплекса Гольджи (КГ), теряют рибосомы и образуют мембранные выступы, от которых отпочковываются мелкие вакуоли, содержащие синтезированные в ЭПС белки. Отделившиеся от ЭПС вакуоли теряют белковую оболочку, сливаются друг с другом и транспортируются с помощью микротрубочек к цис-зоне комплекса Гольджи, где и сливаются с его мембранами.
2.2.3.3. Рибосомы
Рибосома представляет собой элементарную клеточную машину синтеза любых белков клетки. В отличие от других органелл цитоплазмы (пластид, митохондрий, клеточного центра, мембранной вакуолярной системы и др.), рибосомы в клетке многочисленны: за клеточный цикл их образуется 1х107 штук. Рибосомы - это сложные рибонуклеопротеидные частицы, в состав которых входит множество молекул индивидуальных белков и несколько молекул РНК. Р ибосомы прокариот и эукариот по своим размерам отличаются, хотя и обладают общими принципами организации и функционирования. Полная, работающая рибосома, состоит из двух неравных субъединиц: большой и малой. Размер полной прокариотической рибосомы составляет 20 х 17 х 17 нм, эукариотической - 25 х 20 х 20. Полная прокариотическая рибосома имеет коэффициент седиментации 70S и диссоциирует на две субъединицы: 50S и 30S. Полная эукариотическая рибосома, 80S рибосома, диссоциирует на 60S и 40S субъединицы. Малая рибосомная субъединица имеет палочковидную форму с несколькими небольшими выступами. Большая субъединица похожа на полусферу с тремя торчащими выступами. При образовании полной рибосомы, малая субчастица ложится одним концом на один из выступов большой субъединицы, а другим в ее желобок. В состав малых субъединиц входит по одной молекуле РНК, а в состав большой – несколько: у прокариот – две, а у эукариот – 3 молекулы.
ибосомы прокариот и эукариот по своим размерам отличаются, хотя и обладают общими принципами организации и функционирования. Полная, работающая рибосома, состоит из двух неравных субъединиц: большой и малой. Размер полной прокариотической рибосомы составляет 20 х 17 х 17 нм, эукариотической - 25 х 20 х 20. Полная прокариотическая рибосома имеет коэффициент седиментации 70S и диссоциирует на две субъединицы: 50S и 30S. Полная эукариотическая рибосома, 80S рибосома, диссоциирует на 60S и 40S субъединицы. Малая рибосомная субъединица имеет палочковидную форму с несколькими небольшими выступами. Большая субъединица похожа на полусферу с тремя торчащими выступами. При образовании полной рибосомы, малая субчастица ложится одним концом на один из выступов большой субъединицы, а другим в ее желобок. В состав малых субъединиц входит по одной молекуле РНК, а в состав большой – несколько: у прокариот – две, а у эукариот – 3 молекулы.
Рибосомные РНК обладают сложной вторичной и третичной структурой, образуя сложные петли и шпильки на комплементарных участках, что приводит к самоупаковке, самоорганизации этих молекул в сложное по форме тело.
2.2.3.4. Комплекс Гольджи (КГ)
В 1898 году итальянский ученый Камилло Гольджи, используя свойства связывания тяжелых металлов (осмия и серебра) с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал «внутренним сетчатым аппаратом». Обычно элементы комплекса Гольджи расположены около ядра, вблизи клеточного центра. Участки комплеса Гольджи, имеют в некоторых клетках вид сложных сетей, где ячейки связаны друг с другом или представлены в виде отдельных темных участков, лежащих независимо друг от друга, имеющих вид палочек, зерен, вогнутых дисков. В электронном микроскопе видно, что комплекс Гольджи представлен мембранными структурами, собранными вместе в небольшой зоне. Отдельная зона скопления этих мембран называется диктиосомой. В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены в виде стопки плоские мембранные мешки, или цистерны, между которыми располагаются тонкие прослойки гиалоплазмы. Каждая отдельная цистерна имеет диаметр около 1 мкм; в центре ее мембраны могут быть сближены (25 нм), а на периферии имеют расширения, ампулы. Количество таких мешков в стопке обычно не превышает 5-10. Кроме плотно расположенных плоских цистерн в зоне комплекса Гольджи наблюдается множество вакуолей. Мелкие вакуоли встречаются главным образом в периферических участках; иногда видно, как они отшнуровываются от ампулярных расширений на краях плоских цистерн.
1898 году итальянский ученый Камилло Гольджи, используя свойства связывания тяжелых металлов (осмия и серебра) с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал «внутренним сетчатым аппаратом». Обычно элементы комплекса Гольджи расположены около ядра, вблизи клеточного центра. Участки комплеса Гольджи, имеют в некоторых клетках вид сложных сетей, где ячейки связаны друг с другом или представлены в виде отдельных темных участков, лежащих независимо друг от друга, имеющих вид палочек, зерен, вогнутых дисков. В электронном микроскопе видно, что комплекс Гольджи представлен мембранными структурами, собранными вместе в небольшой зоне. Отдельная зона скопления этих мембран называется диктиосомой. В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены в виде стопки плоские мембранные мешки, или цистерны, между которыми располагаются тонкие прослойки гиалоплазмы. Каждая отдельная цистерна имеет диаметр около 1 мкм; в центре ее мембраны могут быть сближены (25 нм), а на периферии имеют расширения, ампулы. Количество таких мешков в стопке обычно не превышает 5-10. Кроме плотно расположенных плоских цистерн в зоне комплекса Гольджи наблюдается множество вакуолей. Мелкие вакуоли встречаются главным образом в периферических участках; иногда видно, как они отшнуровываются от ампулярных расширений на краях плоских цистерн.
Принято различать в зоне диктиосомы проксимальный или формирующийся, цис-участок, и дистальный или зрелый, транс-участок. Между ними располагается средний или промежуточный участок комплекса Гольджи.
В секретирующих клетках обычно комплекс Гольджи поляризован: его проксимальная часть обращена к цитоплазме и ядру, а дистальная - к поверхности клетки.
К омплекс Гольджи является промежуточным звеном между собственно синтезом секретируемого белка и выведением его из клетки. Синтезированный на рибосомах экспортируемый белок отделяется и накапливается внутри цистерн ЭПС, по которым он транспортируется к зоне мембран комплекса Гольджи. Здесь от гладких участков ЭПС отщепляются мелкие вакуоли, содержащие синтезированный белок, которые поступают в зону вакуолей в проксимальной части диктиосомы. В этом месте вакуоли могут сливаться друг с другом и с плоскими цис-цистернами диктиосомы. Происходит перенесение белкового продукта уже внутри полостей цистерн КГ. По мере созревания белков в цистернах комплекса Гольджи, они с помощью мелких вакуолей переносятся от цистерн к цистерне в дистальную часть диктиосомы, пока не достигают трубчатой мембранной сети в транс-участке диктиосомы. В этом участке происходит отщепление мелких пузырьков, содержащих уже зрелый продукт. Отделившиеся мелкие пузырьки сливаются друг с другом, образуя секреторные вакуоли. После этого секреторные вакуоли начинают двигаться к поверхности клетки, соприкасаются с плазматической мембраной, с которой сливаются их мембраны, и, таким образом, содержимое этих вакуолей оказывается за пределами клетки. Этот процесс напоминает пиноцитоз, только с обратной последовательностью стадий, и называется экзоцитоз.
омплекс Гольджи является промежуточным звеном между собственно синтезом секретируемого белка и выведением его из клетки. Синтезированный на рибосомах экспортируемый белок отделяется и накапливается внутри цистерн ЭПС, по которым он транспортируется к зоне мембран комплекса Гольджи. Здесь от гладких участков ЭПС отщепляются мелкие вакуоли, содержащие синтезированный белок, которые поступают в зону вакуолей в проксимальной части диктиосомы. В этом месте вакуоли могут сливаться друг с другом и с плоскими цис-цистернами диктиосомы. Происходит перенесение белкового продукта уже внутри полостей цистерн КГ. По мере созревания белков в цистернах комплекса Гольджи, они с помощью мелких вакуолей переносятся от цистерн к цистерне в дистальную часть диктиосомы, пока не достигают трубчатой мембранной сети в транс-участке диктиосомы. В этом участке происходит отщепление мелких пузырьков, содержащих уже зрелый продукт. Отделившиеся мелкие пузырьки сливаются друг с другом, образуя секреторные вакуоли. После этого секреторные вакуоли начинают двигаться к поверхности клетки, соприкасаются с плазматической мембраной, с которой сливаются их мембраны, и, таким образом, содержимое этих вакуолей оказывается за пределами клетки. Этот процесс напоминает пиноцитоз, только с обратной последовательностью стадий, и называется экзоцитоз.
В комплексе Гольджи происходит не просто «перекачка» продуктов из одной полости в другую, но и постепенно идет их «созревание», модификация белков, которая заканчивается «сортировкой» продуктов, направляющихся или к лизосомам, или к плазматической мембране, или к секреторным вакуолям.
комплексе Гольджи происходит не просто «перекачка» продуктов из одной полости в другую, но и постепенно идет их «созревание», модификация белков, которая заканчивается «сортировкой» продуктов, направляющихся или к лизосомам, или к плазматической мембране, или к секреторным вакуолям.
2.2.3.5. Лизосомы
Лизосомы не представляют собой в клетках самостоятельных структур, они образуются за счет активности ЭПС и комплекса Гольджи, они участвуют в процессах внутриклеточного расщепления биологических макромолекул.
Лизосомы были открыты Де Дюв в 1955году. Лизосомы представляют собой окруженные липопротеидной мембраной пузырьки, содержащие около 40 гидролитических ферментов: протеиназы, нуклеазы, гликозидазы, фосфорилазы, фосфатазы, сульфитазы и др. Под электронным микроскопом видно, что лизосомы представлены пузырьками (0,2-0,4 мкм), ограниченными одиночной мембраной, с очень разнородным содержимым внутри. Во фракции лизосом встречаются пузырьки с гомогенным, бесструктурным содержимым, и пузырьки, заполненные плотным веществом, содержащим в свою очередь вакуоли; часто можно видеть внутри лизосом не только участки мембран, но и фрагменты митохондрий и ЭПС.
Было обнаружено, что среди различных по морфологии лизосом можно выделить четыре типа: первичные лизосомы, вторичные лизосомы, аутофагосомы и остаточные тельца. Первичные лизосомы представляют собой мелкие мембранные пузырьки размером около 100 нм, заполненные бесструктурным веществом, содержащим набор гидролаз и в том числе кислую фосфатазу, - характерный для лизосом фермент. Эти мелкие вакуоли, первичные лизосомы, трудно отличить от мелких вакуолей на периферии зоны комплекса Гольджи. С помощью ряда точных экспериментов установили, что в дальнейшем первичные лизосомы сливаются с фагоцитарными или пиноцитозными вакуолями, эндосомами, образуя вторичную лизосому или внутриклеточную пищеварительную вакуоль. При этом содержимое первичной лизосомы сливается с полостью эндоцитозной вакуоли, и гидролазы первичной лизосомы получают доступ к веществам, которые они и начинают расщеплять.
Разнообразие по величине и по структуре клеточных лизосом связано в первую очередь с разнообразием вторичных лизосом - продуктов слияния эндоцитозных вакуолей с первичными лизосомами. Таким образом, вторичные лизосомы представляют собой не что иное, как внутриклеточные пищеварительные вакуоли, ферменты которых доставлены с помощью мелких первичных лизосом. Поэтому от типа поглощенных веществ или частичек зависит размер и внутренняя структура таких лизосом.
Р асщепление, переваривание биогенных макромолекул внутри лизосом может идти в ряде клеток не до конца. В этом случае в полостях лизосом происходит накопление непереваренных продуктов, происходит переход вторичных лизосом в телолизосомы, или остаточные тельца. Часто в остаточных тельцах наблюдается вторичная структуризация непереваренных липидов, которые образуют сложные слоистые структуры. Там же происходит отложение пигментных веществ. У человека при старении организма в клетках мозга, печени и в мышечных волокнах в телолизосомах происходит отложение «пигменте старения» - липофусцина.
асщепление, переваривание биогенных макромолекул внутри лизосом может идти в ряде клеток не до конца. В этом случае в полостях лизосом происходит накопление непереваренных продуктов, происходит переход вторичных лизосом в телолизосомы, или остаточные тельца. Часто в остаточных тельцах наблюдается вторичная структуризация непереваренных липидов, которые образуют сложные слоистые структуры. Там же происходит отложение пигментных веществ. У человека при старении организма в клетках мозга, печени и в мышечных волокнах в телолизосомах происходит отложение «пигменте старения» - липофусцина.
Аутолизосомы (аутофагосомы) постоянно встречаются в клетках простейших, растений и животных. По своей морфологии их относят к вторичным лизосомам, но с тем отличием, что в составе этих вакуолей встречаются фрагменты или даже целые цитоплазматические структуры, такие, как митохондрии, пластиды, элементы ЭПС, рибосомы, гранулы гликогена и т.д. Процесс образования аутофагосом еще недостаточно ясен. По одним представлениям, первичные лизосомы могут выстраиваться вокруг клеточной органеллы, сливаться друг с другом и таким образом отделять ее от соседних участков цитоплазмы - участок оказывается отделенным мембраной и заключенным внутри такой сложной лизосомы. Процесс аутофагоцитоза связан с отбором и уничтожением измененных, «сломанных» клеточных компонентов. В этом случае лизосомы выполняют роль внутриклеточных чистильщиков, контролирующих дефектные структуры. Значительно возрастает число аутофагосом при различных повреждениях клеток.
2.2.3.6. Агланулярная (гладкая) эндоплазматическая сеть
Агранулярная ЭПС представляет собой часть мембранной вакуолярной системы клетки, представлена мембранами, образующими мелкие вакуоли и трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярной, на мембранах гладкой ЭПС нет рибосом. Диаметр вакуолей и канальцев гладкой ЭПС обычно около 50-100 нм. Выраженность мембран гладкой ЭПС может быть неодинаковой как для различных клеток, так и внутри одной клетки. В клетках эпителия кишечника гладкая ЭПС локализуется главным образом в апикальной, верхней части клетки, вблизи всасывающей поверхности. В клетках печени зоны гладкой ЭПС часто связаны с местами отложения гликогена.
Неоднократно была установлена непрерывность перехода между гладкой ЭПС и гранулярной. Цистерны гранулярной ЭПС теряют на своей поверхности рибосомы и становятся гладкими. При этом такой участок цистерны делается неровным, начинает ветвиться, переходя в трубочки и канальцы агранулярной ЭПС. Гладкая ЭПС является вторичной по отношению к гранулярной и происходит из последней.
Деятельность гладкой ЭПС связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов.
Агранулярная эндоплазматическая сеть особенно развита в клетках, секретирующих стероиды, например, в клетках коркового вещества надпочечника, интерстициальных клетках семенников, участвующих в синтезе стероидных гормонов, а также в клетках сальных желез.
В печени часто увеличение зон гладкой ЭПС связано с рядом патологических процессов в клетках. Так, при барбитуратных отравлениях, при действии различных канцерогенов или ядовитых веществ, при действии больших доз гормональных препаратов клетки печени теряют характерную для них базофилию цитоплазмы, в них падает содержание РНК и появляются в цитоплазме оксифильные зоны. В электронном микроскопе эти зоны представлены скоплениями мембран гладкой ЭПС. Это явление связано с тем, что в этих местах происходят процессы разрушения различных вредных веществ.
В поперечнополосатых мышцах каналы гладкой ЭПС окружают каждую миофибриллу. Здесь ЭПС выполняет специальную функцию депонирования ионов кальция.
Среди высших растений агранулярная эндоплазматическая сеть встречается в клетках тканей, участвующих в синтезе и транспорте терпенов, стероидов, липидов.
45
Просмотр содержимого документа
«клетка модуль 4»
2.2.3.7. Вакуоли растительных клеток
Клетки как низших, так и высших растительных организмов содержат в цитоплазме вакуоли, несущие ряд важных физиологических нагрузок. У молодых клеток может быть несколько мелких вакуолей, которые по мере роста и дифференцировки клетки сливаются друг с другом и образуют одну или несколько крупных вакуолей, занимающих до 90% объема всей клетки. Центральные вакуоли отделены от цитоплазмы одинарной мембраной, сходной по толщине с плазмалеммой. Мембрана, ограничивающая центральные вакуоли, носит название тонопласта. Возникают центральные вакуоли из мелких пузырьков, отщепившихся от аппарата Гольджи. Такие первичные вакуоли растут в объеме, сливаются друг с другом и образуют одну или несколько крупных вакуолей, оттесняющих цитоплазму с ядром и органоидами к периферии клетки. Полость вакуоли заполнена клеточным соком, представляющим собой водный раствор, в который входят различные неорганические соли, сахара, органические кислоты и их соли и другие низкомолекулярные соединения, а также некоторые высокомолекулярные вещества (например, белки).
Центральные вакуоли растений выполняют многообразные и важные функции. Одной из главных ее функций является поддержание тургорного давления клеток. Растворенные в соке вакуолей молекулы определяют его осмотическую концентрацию. Другая функция определяется тем, что вакуоль представляет собой большую полость, отделенную от гиалоплазмы мембраной, тонопластом, обладающим свойствами полупроницаемости и через который может происходить, как и через плазматическую мембрану, активный транспорт различных молекул. Поэтому вакуоли могут использоваться клетками как накопительные резервуары не только для отложения запасных веществ, но и для выброса метаболитов, для экскреции. В вакуолях происходит отложение многих глюкозидов, к которым относятся различные пигменты, например, антоцианы.
Другой обширный ряд функций вакуолей связан с накоплением запасных веществ, таких как сахара и белки. Сахара в вакуолях содержатся в виде растворов, встречаются и резервные полисахариды типа инулина. Запасание белков семян злаковых происходит в так называемых алейроновых вакуолях, которые заполняются альбуминами и глобулинами, после чего вакуоли обезвоживаются, превращаясь в твердые алейроновые зерна. При прорастании семян эти зерна обводняются и снова превращаются в вакуоли.
2.2.3.8. Сферосомы
Сферосомы - это мембранные пузырьки, встречающиеся в клетках растений. Они окрашиваются липофильными красителями, имеют высокий коэффициент преломления и поэтому хорошо видны в световой микроскоп. Сферосомы образуются из элементов эндоплазматической сети. На конце цистерны ЭПС начинает накапливаться осмиофильный материал, затем от этого участка отшнуровывается и начинает расти мелкий пузырек, достигающий диаметра 0,1-0,5 мкм. Рост сферосом и перестройка их содержимого связаны с накоплением в них масла, так что сферосома постепенно превращается в масляную каплю. Кроме жиров в составе сферосом обнаруживают белки и среди них фермент липазу, расщепляющую липиды.
2.2.3.9. Пероксисомы
Пероксисомы (микротельца) - это небольшие вакуоли (0,3-1,5 мкм), одетые одинарной мембраной, отграничивающей гранулярный матрикс, в центре которого располагается сердцевина. В зоне сердцевины часто, особенно в пероксисомах печеночных клеток, видны кристаллоподобные структуры, состоящие из регулярно упакованных фибрилл или трубочек. Изолированные сердцевины пероксисом содержат фермент уратоксидазу.
Пероксисомы обнаружены у простейших (амебы, тетрахимена), у низших грибов (дрожжи), у высших растений в некоторых эмбриональных тканях (эндосперм) и в зеленых частях, способных к фотореспирации, у высших позвоночных животных они обнаруживаются главным образом в печени и почках. В печени крыс на клетку число пероксисом колеблется от 70 до 100.
Пероксисомы часто локализуются вблизи мембран ЭПС. У зеленых растений пероксисомы не редко находятся в тесном контакте с митохондриями и пластидами.
Впервые пероксисомы были выделены из печени и почек. Во фракциях пероксисом обнаруживаются ферменты, связанные с метаболизмом перекиси водорода. Это ферменты (оксидазы, уратоксидаза, оксидаза d-аминокислот) окислительного дезаминирования аминокислот, при работе которых образуется перекись водорода (Н2О2) и каталаза, разрушающая ее. В пероксисомах печени каталаза составляет до 40 % всех белков и локализована в матриксе. Так как Н2О2 является токсическим веществом для клеток, то каталаза пероксисом может играть важную защитную роль. Пероксисомы цыплят и лягушек кроме уратоксидазы содержат ряд ферментов катаболизма пуринов.
У животных и некоторых растений (проростки клещевины) пероксисомы играют важную роль при превращении жиров в углеводы. В пероксидах происходит накопление специфических белков, которые синтезируются в цитозоле.
2.2.3.10. Митохондрии
Для осуществления любых клеточных функций необходима энергия. Клетки синтезируют молекулы АТФ (аденозинтрифосфат), обладающие высокоэнергетическими фосфатными связями. При разрушении этих связей выделяемая энергия может тратиться на любые клеточные функции: на активный транспорт веществ, на синтетические процессы, на механическую работу и т.д. В клетках животных синтез АТФ осуществляется специальными органеллами - митохондриями, в растительных клетках кроме митохондрий в энергообеспечении огромную роль играют хлоропласты. Митохондрии и пластиды - это двумембранные органеллы эукариотических клеток. Митохондрии присутствуют во всех к летках; их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии в синтезе молекул АТФ. Поэтому митохондрии часто называют энергетическими станциями клетки.
летках; их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии в синтезе молекул АТФ. Поэтому митохондрии часто называют энергетическими станциями клетки.
В
первые митохондрии были обнаружены Р. Альтманом, который описал эти клеточные органеллы в конце XIX века, называя их «биобластами». Митохондрии или хондриосомы (от греч. mitos– нить, chondrion- зернышко, soma- тельце) при световой микроскопии имеют вид гранул или нитей. Толщина митохондрий в различных клетках относительно постоянна (около 0,5 мкм), а длина колеблется, достигая у нитчатых форм до 7-60 мкм. В световом микроскопе на окрашенных препаратах не всегда можно определить реальный размер митохондрий.
Изучая митохондрии в электронном микроскопе, также трудно решить вопрос об их истинной длине, так как на срез попадает только незначительный объем митохондрии. Более того, на срезе одна извитая митохондрия может быть представлена несколькими сечениями, и только пространственная трехмерная реконструкция, построенная на изучении серийных срезов, может решить вопрос, имеем ли мы дело с тремя-шести отдельными митохондриями или же с одной изогнутой или разветвленной. Длинные ветвящиеся митохондрии были описаны в клетках культуры ткани млекопитающих, в клетках многих растений. Обычные же подсчеты показывают, что на печеночную клетку приходится около 200 митохондрий. Это составляет более 20% от общего объема цитоплазмы и около 30-35% от общего количества белка в клетке. П лощадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности ее плазмолеммы. Локализация митохондрий в клетках быть различной. В малодифференцированных клетках меристемы растений митохондрии располагаются более или менее равномерно. В клетках эпителия почечных канальцев митохондрии ориентированы вдоль продольной оси клетки. Обычно митохондрии скапливаются вблизи тех участков цитоплазмы, где возникает потребность в АТФ. Так, в скелетных мышцах митохондрии находятся вблизи миофибрилл, в сперматозоидах образуют спиральный футляр вокруг оси жгутика. В аксонах нервных клеток митохондрии располагаются в синапсах, где происходит процесс передачи нервного импульса. Митохондрии независимо от их величины или формы имеют универсальное строение, их ультраструктура однообразна.
лощадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности ее плазмолеммы. Локализация митохондрий в клетках быть различной. В малодифференцированных клетках меристемы растений митохондрии располагаются более или менее равномерно. В клетках эпителия почечных канальцев митохондрии ориентированы вдоль продольной оси клетки. Обычно митохондрии скапливаются вблизи тех участков цитоплазмы, где возникает потребность в АТФ. Так, в скелетных мышцах митохондрии находятся вблизи миофибрилл, в сперматозоидах образуют спиральный футляр вокруг оси жгутика. В аксонах нервных клеток митохондрии располагаются в синапсах, где происходит процесс передачи нервного импульса. Митохондрии независимо от их величины или формы имеют универсальное строение, их ультраструктура однообразна.
Митохондрии ограничены двумя мембранами. Наружняя митохондриальная мембрана отделяет ее от гиалоплазмы. Обычно она имеет ровные контуры, она представляет собой мембранный мешок. Наружнюю мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм. Внутренняя мембрана ограничивает собственно внутреннее содержимое митохондрии, ее матрикс. Характерной чертой внутренней мембраны митохондрий является их способность образовывать многочисленные впячивания внутрь митохондрий. Такие впячивания чаще всего имеют вид плоских гребней, или крист. Матрикс митохондрий имеет тонкозернистое гомогенное строение, в нем иногда выявляются тонкие собранные в клубок нити (около 2-3 нм) и гранулы около 15-20нм. Теперь стало известно, что нити матрикса митохондрий представляют собой молекулы ДНК в составе митохондриального нуклеоида, а мелкие гранулы - митохондриальные рибосомы. Хондриом – это совокупность всех митохондрий в одной клетке.
2.2.3.11. Пластиды
Пластиды – это мембранные органоиды, встречающиеся у фотосинтезирующих эукариотических организмов (высшие растения, низшие водоросли, некоторые одноклеточные организмы). Подобно митохондриям, пластиды окружены двумя мембранами, в их матриксе имеется собственная геномная система, функции пластид связаны с энергообеспечением клетки, идущим на нужды фотосинтеза. У высших растений найден целый набор различных пластид (хлоропласт, лейкопласт, амилопласт, хромопласт), представляющих собой ряд взаимных превращений одного вида пластиды в другой. Основной структурой, которая осуществляет фотосинтетические процессы, является хлоропласт.
Хлоропласты – двумембранные органеллы дисковидной формы с шириной 2-4 мкм и протяженностью 5-10 мкм. У зеленых водорослей встречаются гигантские хлоропласты (хроматофоры), достигающие длины 50 мкм. Количество хлоропластов в клетках разных растений не одинаково. Так, у зеленых водорослей может быть по одному хлоропласту на клетку. Обычно на клетку высших растений приходится в среднем 10-30 хлоропластов. Встречаются клетки с большим количеством хлоропластов. Внешняя мембрана, как и внутренняя, имеет толщину около 7 нм, они отделены друг от друга межмембранным пространством около 20-30 нм. Внутренняя мембрана хлоропластов отделяет строму пластиды, аналогичную матриксу митохондрий. В строме зрелого хлоропласта высших растений видны два типа внутренних мембран. Это мембраны, образующие плоские, протяженные ламеллы стромы, и мембраны тилакоидов, плоских дисковидных вакуолей или мешков.
Л амеллы стромы (толщиной около 20 мкм) представляют собой плоские полые мешочки или имеют вид сети из разветвленных и связанных друг с другом каналов, располагающихся в одной плоскости. Обычно ламеллы стромы внутри хлоропласта лежат параллельно друг другу и не образуют связей между собой.
амеллы стромы (толщиной около 20 мкм) представляют собой плоские полые мешочки или имеют вид сети из разветвленных и связанных друг с другом каналов, располагающихся в одной плоскости. Обычно ламеллы стромы внутри хлоропласта лежат параллельно друг другу и не образуют связей между собой.
К Рис. 38. Строение хлоропласта (а), лейкопласта (б), хромопласта (в) 1 – наружная мембрана; 2 – внут-ренняя мембрана; 3 – матрикс; 4 – лямеллы стромы; 5 – грана; 6 – тилакоид; 7 – крахмальное зерно; 8 – липидная капля с пигментами амелл стромы. Ламеллы стромы и мембраны тилакоидов образуются путем отделения от внутренней мембраны при начальных этапах развития пластид.
амелл стромы. Ламеллы стромы и мембраны тилакоидов образуются путем отделения от внутренней мембраны при начальных этапах развития пластид.
В матриксе (строме) хлоропластов обнаруживаются молекулы ДНК, рибосомы; там же происходит первичное отложение запасного полисахарида крахмала в виде крахмальных зерен.
Хлоропласты - это структуры, в которых происходят фотосинтетические процессы, приводящие к связыванию углекислоты, синтезу сахаров и выделению кислорода.
Характерным для хлоропластов является наличие в них пигментов, хлорофиллов, которые и придают окраску зеленым растениям. При помощи хлорофилла зеленые растения поглощают энергию солнечного света и превращают ее в химическую. Поглощение света с определенной длиной волны приводит к изменению в структуре молекулы хлорофилла, она переходит при этом в возбужденное, активированное состояние. Освобождающаяся энергия активированного хлорофилла приводит к синтезу АТФ, энергия которой тратится на реакции синтеза сахаров.
Процесс развития различных пластид можно представить в виде идущего в одном направлении ряда смены форм:
Пропластида лейкопласт хлоропласт хромопластамилопласт.
Многими исследованиями был установлен необратимый характер онтогенетических переходов пластид. У высших растений возникновение и развитие хлоропластов происходят через изменения пропластид. Пропластиды представляют собой мелкие (0,4-1 мкм) двумембранные пузырьки, не имеющие отличительных черт их внутреннего строения. Они отличаются от вакуолей цитоплазмы более плотным содержимым и наличием двух отграничивающих мембран. Внутренняя мембрана может давать небольшие складки или образовывать мелкие вакуоли. Пропластиды чаще всего встречаются в делящихся тканях растений. Судьба таких пропластид будет зависеть от условий развития растений. При нормальном освещении пропластиды превращаются в хлоропласты. Сначала они растут, при этом происходит образование продольно расположенных мембранных складок от внутренней мембраны. Одни из них простираются по всей длине пластиды и формируют ламеллы стромы; другие образуют ламеллы тилакоидов, которые выстраиваются в виде стопки и образуют граны зрелых хлоропластов.
Лейкопласты отличаются от хлоропластов отсутствием развитой ламеллярной системы. Встречаются они в клетках запасающих тканей. Из-за их неопределенной морфологии лейкопласты трудно отличить от пропластид, а иногда и от митохондрий. Они, как и пропластиды, бедны ламеллами, но, тем не менее, способны к образованию под влиянием света нормальных тилакоидных структур и к приобретению зеленой окраски. В темноте лейкопласты могут накапливать различные запасные вещества, а в строме лейкопластов откладываются зерна вторичного крахмала. Другой формой пластид у высших растений является хромопласт, окрашивающийся обычно в желтый свет в результате накопления в нем каротиноидов. Хромопласты образуются из хлоропластов и значительно реже их лейкопластов (например, в корне моркови).
2.2.3.12. Клеточный центр и центриоли
Клеточный центр (центросомы) был обнаружен и описан более ста лет назад (Флемминг, 1875; Э. Бенеден, 1876). Клеточный центр характерен для клеток животных и низших растений. Отсутствует клеточный центр у высших растений, у низших грибов и некоторых простейших. Обычно эта органелла располагается в геометрическом центре клетки, поэтому и названа клеточным центром. Наиболее типичное строение клеточный центр имеет в клетках животных: он представляет собой зону, состоящую из центриолей, окруженных фибриллярной массой.
Тонкое строение центриолей удалось изучить с помощью электронного микроскопа. Основу строения центриолей составляют расположенные по окружности девять триплетов микротрубочек, образующие полый цилиндр (шириной около 0,15 мкм, а длиной - 0,3-0,5 мкм). Каждый триплет микротрубочек располагается к радиусу цилиндра под углом около 400.
С истему микротрубочек центриоли обычно описывают формулой 9 + 0, или (9х3) + 0, подчеркивая отсутствие микротрубочек в её центральной части.
истему микротрубочек центриоли обычно описывают формулой 9 + 0, или (9х3) + 0, подчеркивая отсутствие микротрубочек в её центральной части.
Вокруг каждой центриоли расположен бесструктурный, или тонковолокнистый матрикс. Около центриолей можно обнаружить несколько дополнительных структур: это сателлиты, фокусы схождения микротрубочек, исчерченные волокнистые корешки и дополнительные микротрубочки, образующие вокруг центриоли особую зону - центросферу.
Обычно в интерфазных клетках присутствуют две центриоли, располагающиеся рядом друг с другом, образуя дуплет центриолей, или диплосому. В диплосоме центриоли располагаются под прямым углом по отношению друг к другу. Из двух центриолей различают «материнскую» и «дочернюю». В дистальном участке материнской центриоли располагается аморфный материал в виде выростов или шпор - это придатки. Их нет на дочерней центриоли. Только материнская центриоль содержит сателлиты, состоящие из конусовидной ножки, расположенной на стенке центриоли, и головки, заканчивающейся на этой ножке. Ножки сателлитов часто имеют поперечную исчерченность. Микротрубочки отходят и от головок сателлитов. Эти центросомные микротрубочки не отходят непосредственно от микротрубочек цилиндров центриолей, а связаны или с сателлитами, или с матриксом. Такие микротрубочки и образуют лучистую сферу - центросферу вокруг центриоли.
Центриоли в делящихся клетках принимают участие в формировании веретена деления и располагаются на полюсах клетки. В неделящихся клетках центросомы часть определяют полярность клеток эпителия и располагаются вблизи аппарата Гольджи.
2.2.4. Ядро эукариотических клеток
Сам термин «ядро» впервые был применен М. Брауном в 1833 г. для обозначения шаровидных постоянных структур в клетках растений. Позднее такую же структуру описали во всех клетках высших организмов. Большинство клеток содержат одно ядро, хотя описаны и многоядерные клетки. Эритроциты млекопитающих и зрелые ситовидные клетки растительных организмов не имеют ядра; часто в популярной литературе безъядерными клетками называют тромбоциты, которые являются не клетками, а фрагментами мегакариоцита.
Клетки печени, поджелудочной железы, нервные клетки имеют ядра шаровидной формы, у миоцитов ядра дисковидные. Клетки гранулоцитов крови имеют сегментированные ядра.И тем не менее, самые непохожие по форме ядра, имеют общий план строения. Клеточное ядро состоит из ядерной оболочки, отделяющей его от цитоплазмы, хроматина, ядрышка, ядерного белкового остова (матрикса) и нуклеоплазмы (или ядерного сока). Эти основные компоненты встречаются практически во всех неделящихся клетках эукариотических одно- или многоклеточных организмов.
Ядро выполняет функцию хранения генетической информации, а также обеспечивает реализацию этой генетической информации через управление синтезом белка.
54
Просмотр содержимого документа
«клетка модуль 5»
2.2.4.1. Ядерная оболочка
Структура, ограничивающая клеточное ядро - ядерная оболочка, характерна для эукариотических клеток. Она разделяет два внутриклеточных компартмента друг от друга, цитоплазму от ядра. Разделение этих структур в пространстве приводит к обособлению процессов синтеза белка и процессов синтеза нуклеиновых кислот, что создает дополнительные, по сравнению с прокариотами, возможности для регуляции генной активности. Активная регуляция транспорта из цитоплазмы в ядро и из ядра в цитоплазму через специальные комплексы пор создает систему избирательного транспорта веществ, делая ядерную оболочку «генными воротами», регулирующими потоки ядерного импорта и экспорта. Кроме того, ядерная оболочка играет большую роль в организации трехмерной структуры интерфазного ядра, элементы ядерной оболочки являются частью ядерного белкового матрикса.
Ядерная оболочка состоит из двух мембран, внешней и внутренней, между которыми располагается перинуклеарное пространство. Внутренняя мембрана ядерной оболочки структурно связана с ламиной - фиброзным периферическим слоем ядерного белкового матрикса. Ядерная оболочка имеет особые ядерные поры, которые образуются за счет многочисленных зон слияния двух ядерных мембран и представляют собой как бы округлые, сквозные перфорации всей ядерной оболочки.
В нешняя мембрана ядерной оболочки, непосредственно контактирующая с цитоплазмой клетки, связана с мембранной системой эндоплазматической сети (ЭПС), на ней обычно располагается большое количество рибосом, как и на мембранах эргастоплазмы. Существуют многочисленные наблюдения о непосредственном переходе внешней ядерной мембраны в систему каналов эндоплазматической сети.
нешняя мембрана ядерной оболочки, непосредственно контактирующая с цитоплазмой клетки, связана с мембранной системой эндоплазматической сети (ЭПС), на ней обычно располагается большое количество рибосом, как и на мембранах эргастоплазмы. Существуют многочисленные наблюдения о непосредственном переходе внешней ядерной мембраны в систему каналов эндоплазматической сети.
Внутренняя мембрана ядерной оболочки рибосом на своей поверхности не имеет, но связана с фиброзным слоем, ядерной ламиной, которая, в свою очередь, заякоревает хроматин на ядерной оболочке.
Фиброзный слой ламины все время перестраивается, особенно в связи с ростом поверхности ядра во время клеточного цикла. Характерные для внутренней ядерной мембраны белки ламины A, C и B относятся к фибриллярным белкам.
Наиболее характерной и бросающейся в глаза структурой в составе ядерной оболочки является ядерная пора. Поры в оболочке образуются за счет двух ядерных мембран в виде округлых сквозных отверстий или перфораций с диаметром около 100 нм. Округлое сквозное отверстие в ядерной оболочке заполнено сложно организованными глобулярными и фибриллярными структурами. Совокупность мембранных перфораций и этих структур называют комплексом пор ядра.
Я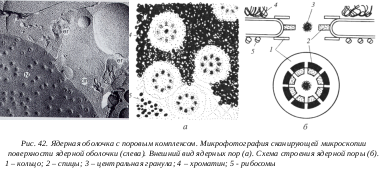 дерный поровый комплекс состоит из более 1000 белков, которые собраны примерно в 12 субкомплексов. Поровый комплекс представляет собой цилиндрическую фигуру. От периферических компонентов порового комплекса в сторону цитоплазмы простираются фибриллы белков. Со стороны ядра тоже фибриллярные выросты образуют корзинкоподобную структуру. Центр цилиндрической фигуры содержит «пробку» (центральную гранулу). Комплекс ядерной поры играет роль в контроле за ядерно-цитоплазменными связями. В процессе ядерно-цитоплазматического транспорта ядерные поры функционируют как некоторое молекулярное сито, пропуская частицы определенного размера пассивно, по градиенту концентрации. Так, ионы, сахара, нуклеотиды, АТФ, гормоны свободно поступают в ядра. С другой стороны ядерные поры осуществляют избирательный транспорт. Так, например, концентрация белков-гистонов в ядре значительно выше, чем в цитоплазме. Но, несмотря на это, во время синтеза ДНК происходит транспорт огромного количества (106 молекул каждые три минуты, или по 100-500 молекул через одну пору за 1 минуту) гистонов из цитоплазмы в ядро.
дерный поровый комплекс состоит из более 1000 белков, которые собраны примерно в 12 субкомплексов. Поровый комплекс представляет собой цилиндрическую фигуру. От периферических компонентов порового комплекса в сторону цитоплазмы простираются фибриллы белков. Со стороны ядра тоже фибриллярные выросты образуют корзинкоподобную структуру. Центр цилиндрической фигуры содержит «пробку» (центральную гранулу). Комплекс ядерной поры играет роль в контроле за ядерно-цитоплазменными связями. В процессе ядерно-цитоплазматического транспорта ядерные поры функционируют как некоторое молекулярное сито, пропуская частицы определенного размера пассивно, по градиенту концентрации. Так, ионы, сахара, нуклеотиды, АТФ, гормоны свободно поступают в ядра. С другой стороны ядерные поры осуществляют избирательный транспорт. Так, например, концентрация белков-гистонов в ядре значительно выше, чем в цитоплазме. Но, несмотря на это, во время синтеза ДНК происходит транспорт огромного количества (106 молекул каждые три минуты, или по 100-500 молекул через одну пору за 1 минуту) гистонов из цитоплазмы в ядро.
Из ядра через поры выходят в цитоплазму субъединицы рибосом и другие рибонуклеопротеиды. Эти данные показывают, что комплексы ядерных пор не представляют собой просто механические сита, которые ограничивают транспорт молекул в зависимости от их размеров.
2.2.4.2. Хроматин
Главный компонент ядер – это хроматин, выполняющий генетическую функцию клетки. В хроматиновой ДНК заложена практически вся генетическая информация. После окраски внутри ядра выявляются зоны плотного вещества, которое хорошо воспринимает разные красители, особенно основные. Благодаря такой способности хорошо окрашиваться этот компонент ядра и получил название «хроматин». Способность хроматина воспринимать основные красители определяется тем, что в состав хроматина входит ДНК в комплексе с белками.
Хроматин интерфазных ядер представляет собой несущие ДНК тельца, которые теряют в это время свою компактную форму, разрыхляются, деконденсируются. Степень такой деконденсации хромосом может быть различной в ядрах разных клеток. Когда хромосома или ее участок полностью деконденсирован, тогда эти зоны называют диффузным хроматином. При неполном разрыхлении хромосом в интерфазном ядре видны участки конденсированного хроматина.
С
тепень деконденсации хроматина в интерфазе может отражать функциональную активность этой структуры: чем более диффузен хроматин интерфазного ядра, тем выше в нем синтетические процессы транскрипции и репликации. Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде телец - хромосом, которые выполняют функцию распределения и переноса генетического материала в дочерние клетки.
Е ще в начале 30-х годов XX века было замечено Э. Гейтцем, что в интерфазных ядрах существуют постоянные участки конденсированного хроматина, наличие которого не зависит от степени дифференцированнности ткани или от функциональной активности клеток. Такие участки получили название гетерохроматина, в отличие от остальной массы хроматина - эухроматина (собственно хроматина). Первоначально понятие гетерохроматина имело сугубо морфологическое значение, потому что при изучении препаратов окрашенных ядер нельзя знать, может ли данный участок конденсированного хроматина перейти в разрыхленное, эухроматическое состояние. На самом же деле только постоянно конденсированные участки хромосом в интерфазных ядрах сейчас принято называть гетерохроматином. Гетерохроматин генетически не активен, он не транскрибируется, реплицируется он позже всего остального хроматина. В состав гетерохроматина входит особая ДНК, обогащенная высокоповторяющимися последовательностями нуклеотидов; он локализован в центромерных, теломерных и интеркалярных зонах митотических хромосом. Функциональное значение гетерохроматина до конца не выяснено, предполагается, что он участвует в конъюгации хромосом во время мейоза.
ще в начале 30-х годов XX века было замечено Э. Гейтцем, что в интерфазных ядрах существуют постоянные участки конденсированного хроматина, наличие которого не зависит от степени дифференцированнности ткани или от функциональной активности клеток. Такие участки получили название гетерохроматина, в отличие от остальной массы хроматина - эухроматина (собственно хроматина). Первоначально понятие гетерохроматина имело сугубо морфологическое значение, потому что при изучении препаратов окрашенных ядер нельзя знать, может ли данный участок конденсированного хроматина перейти в разрыхленное, эухроматическое состояние. На самом же деле только постоянно конденсированные участки хромосом в интерфазных ядрах сейчас принято называть гетерохроматином. Гетерохроматин генетически не активен, он не транскрибируется, реплицируется он позже всего остального хроматина. В состав гетерохроматина входит особая ДНК, обогащенная высокоповторяющимися последовательностями нуклеотидов; он локализован в центромерных, теломерных и интеркалярных зонах митотических хромосом. Функциональное значение гетерохроматина до конца не выяснено, предполагается, что он участвует в конъюгации хромосом во время мейоза.
Основная масса хроматина ядра может менять степень своей компактизации в зависимости от функциональной активности, она относится к эухроматину. Эухроматические неактивные участки, которые находятся в конденсированном состоянии, стали называть факультативным гетерохроматином, подчеркивая необязательность такого его состояния. Хорошим примером факультативного гетерохроматина может служить X-хромосома в организме человека. В клетках мужской особи X-хромосома деконденсирована, она активна, транскрибируется и морфологически не выявляется из-за своего рыхлого, диффузного состояния. В клетках женского организма, где присутствуют две X-хромосомы, одна из них находится в активном, диффузном состоянии, а вторая - в неактивном, конденсированном, она временно гетерохроматизована. В этом состоянии она может существовать в течение всей жизни организма. Но потомки ее, попадая в клетки мужского организма следующего поколения, снова будут активированы.
В дифференцированных клетках всего лишь около 10% генов находится в активном состоянии, остальные гены инактивированы и, соответственно, находятся в составе конденсированного хроматина (факультативный гетерохроматин). Это обстоятельство объясняет, почему большая часть хроматина ядра структурирована.
2.2.4.3. Ядрышко
Внутри интерфазных ядер на окрашенных препаратах видны мелкие, обычно шаровидные тельца - ядрышки. Впервые ядрышки были описаны Фонтана в 1774 г. Ядрышки обнаруживаются практически во всех ядрах эукариотических клеток. В клеточном цикле ядрышко присутствует в течение всей интерфазы: в профазе оно постепенно исчезает, и отсутствует в мета- и анафазе и вновь появляется в середине телофазы.
В озникновение ядрышек связано с определенными зонами на особых, ядрышкообразующих участках хромосом. Эти зоны были названы ядрышковыми организаторами, а сами ядрышки представлялись как структурное выражение хромосомной активности.
озникновение ядрышек связано с определенными зонами на особых, ядрышкообразующих участках хромосом. Эти зоны были названы ядрышковыми организаторами, а сами ядрышки представлялись как структурное выражение хромосомной активности.
Уже в 50-х годах XX века при изучении ультраструктуры ядрышек в их составе были обнаружены гранулы, сходные по своим свойствам с цитоплазматическими гранулами рибонуклеопротеидной природы, с рибосомами. Следующим этапом в изучении ядрышка было открытие принципиального факта: «ядрышковый организатор» является вместилищем генов рибосомных РНК.
Внутренняя часть ядрышка имеет фибриллярную структуру, состоящую из участка ДНК – ядрышкового организатора и синтезирующихся нитей рРНК. Наружная гранулярная часть - это зона зрелых и созревающих субъединиц рибосом. Ядрышко участвует в образовании рибосом и мощно развито в клетках с интенсивным синтезом белка.
Ядерный матрикс состоит из трех морфологических компонентов: периферического белкового сетчатого слоя - лямины, внутреннего, или интерхроматинового матрикса, и «остаточного» ядрышка. Лямина образует сплошной фибриллярный слой, прилежащий к внутренней мембране кариолеммы, поддерживает морфологическую целостность ядра. Интерхроматиновый матрикс представлен рыхлой фибриллярной сетью, в ячейках которых располагается хроматин в разной стадии конденсации. К фибриллам присоединены комплексы ферментов синтеза нуклеиновых кислот. Остаточное ядрышко состоит из плотно уложенных фибрилл и повторяет форму ядрышка. Компоненты ядерного матрикса динамичны и меняются в зависимости от функциональных особенностей клеток.
РЕКОМЕНДАЦИИ ДЛЯ САМОПОДГОТОВКИ
При подготовке к практическому занятию по материалам учебного пособия продумайте, с участием каких органелл в клетке происходят следующие процессы:
| Процессы | Органеллы, клеточные структуры | Особенности ультраструктуры |
| Синтез липидов, холестерина | | |
| Синтез гликогена, полисахаридов | |
|
| Детоксикация ядов | |
|
| Транспорт веществ в клетке | |
|
| Синтез белков | |
|
| Формирование третичной и четвертичной структуры белка | | |
| Присоединение сахаров к белкам | |
|
| Образование лизосом | |
|
| Образование клеточных мембран | |
|
| Внутриклеточное пищеварение | |
|
| Процессы биологического окисления | |
|
| Хранение генетической информации | |
|
| Аккумуляция энергии солнца | |
|
| Образование рибосом | |
|
| Процессы транскрипции | |
|
| Процессы трансляции | |
|
| Обезврещивание Н2О2 | |
|
| Экзо – и эндоцитоз | |
|
| Формирование псевдоподий | |
|
| Передача нервного импульса с одного нейрона на другой | |
|
59
Просмотр содержимого документа
«клетка модуль 6»
тест для проверки знаний
Выберите номер правильного ответа1. Грибы нельзя относить к царству растений, т.к. в их клетках отсутствуют
оболочки
хлоропласты
ядра
митохондрии
2. Гетеротрофный способ питания, отсутствие плотной оболочки и хлоропластов, наличие ядра в клетке – признаки организма царства
животных
грибов
бактерий
растений
3. Изучением строения и функций клетки занимается наука
эмбриология
генетика
селекция
цитология
4. «Сходство обмена веществ в клетках организмов всех царств живой природы» - это положение теории
хромосомной
клеточной
эволюционной
происхождения жизни
5. О единстве органического мира свидетельствует
круговорот веществ
клеточное строение организмов
взаимосвязь организма и среды
приспособленность организмов к среде
6. Для изучения места расположения органоидов в клетке используют метод
микроскопии
центрифугирования
эксперимента
выращивания клеток
7. В клетках человека и животных в качестве строительного материала и источника энергии используются
гормоны и витамина
вода и углекислый газ
неорганические вещества
белки, жиры и углеводы
8. Бактерии не имеют оформленного ядра, поэтому их относят к группе организмов
эукариот
прокариот
автотрофов
гетеротрофов
9. В клетках гетеротрофов в отличие от автотрофов отсутствуют
митохондрии
лизосомы
хлоропласты
рибосомы
10. Внутренняя полужидкая среда клетки, пронизанная мельчайшими нитями и трубочками, в которой расположены органоиды и ядро - это
вакуоль
цитоплазма
комплекс Гольджи
митохондрии
11. Органоиды, состоящие из особого вида нуклеиновых кислот, расположенные на гранулярной эндоплазматической сети и участвующие в биосинтезе белка, - это
митохондрии
лизосомы
рибосомы
хлоропласты
12. Органоид, ограниченный от цитоплазмы одной мембраной, содержащий множество ферментов, которые расщепляют сложные органические вещества до простых, мономеров –
митохондрии
аппарат Гольджи
рибосома
лизосома
13. Отрезок молекулы ДНК, содержащий информацию о первичной структуре белка, называют
геном
генотипом
генетическим кодом
триплетом
14. Из одной молекулы нуклеиновой кислоты в соединении с белками состоит
митохондрия
хлоропласт
хромосома
эндоплазматическая сеть
15. В клетках всех организмов происходят процессы питания, дыхания, поэтому ее называют единицей
строения
генетической
размножения
жизнедеятельности
16. Всю совокупность химических реакций в клетке называют
фотосинтезом
хемосинтезом
брожением
метаболизмом
17. Ускоряют химические реакции в клетке
ферменты
пигменты
витамины
гомоны
18. Клетка – единица развития всех организмов, так как
все клетки имеют одинаковый химический состав
из клеток состоят ткани и органы
клетки имеют сходное строение
благодаря делению и дифференциации клеток формируются ткани
19. Эукариотические животные клетки отличает
отсутствие изолированного ядра
наличие локализованного ядра
наличие локализованного ядра и митохондрий
наличие локализованного ядра, митохондрий и центриолей.
Ионы в клетку поступают
с помощью ферментов клеточной оболочки
пассивным путем ( разница концентрации)
с помощью пиноцитоза
с помощью ферментов под действием разницы концентраций
21. Крупные органические молекулы в клетку поступают
с помощью ферментов клеточной оболочки
пассивным путем (разница концентрации)
с помощью пиноцитоза
с помощью ферментов под действием разницы концентраций
22. Сборка гемоглобина в клетках происходит
ЭПС
митохондриях
аппарате Гольджи
лизосомах
2 3. На рисунке изображено
3. На рисунке изображено
клеточный центр
комплекс Гольджи
хлоропласт
митохондрия
24 На рисунке изображена молекула.
Б
NH2 – CH – COOH
I
CH2
I
SH
елкаДНК
Аминокислоты
Клетчатки
25 На рисунке изображена рибосома, об этом свидетельствуют входящие в её состав
м
 ногочисленные кристы
ногочисленные кристысистемы гран
цистерны и полости
большая и малая субъединицы
2 
вакуолях
цитоплазме
эндоплазматической сети
комплексе Гольджи

2 7. На рисунке изображена молекула, входящая в состав многих структур клетки. Это молекула
7. На рисунке изображена молекула, входящая в состав многих структур клетки. Это молекула
белка
ДНК
углевода
липида
28 На рисунке изображён хлоропласт, который можно узнать по наличию в нём
к
 рист
ристполостей и цистерн
гран
ядрышек
29 Функцию синтеза белков выполняет
гладкая ЭПС
рибосомы на гранулярной ЭПС
митохондрии
лизосомы
аппарат Гольджи
30. Функцию синтеза АТФ выполняет
гладкая ЭПС
рибосомы на гранулярной ЭПС
митохондрии
лизосомы
аппарат Гольджи
31 Функцию сборки и дозревания секрета выполняет
гладкая ЭПС
рибосомы на гранулярной ЭПС
митохондрии
лизосомы
аппарат Гольджи
У
ФУНКЦИИ
становить соответствие32. КЛЕТОЧНЫЕ СТРУКТУРЫ

А. ядро Б. ядрышко В. Фагоцитарная вакуоль Г. Лизосома Д. Тонопласт Е. Рибосомы Ж.Клеточный центр З. Гладкая ЭПС И. Включения К. Вырост цитоплазмы Л. Митохондрии М.Гранулярная ЭПС Н.Комплекс Гольджи
Ответы: 1...2...3...4.....5....6....7….8….9….10……11…….12…….
рекомендации К САМОСТОЯТЕЛЬНОЙ ПРАКТИЧЕСКОЙ РАБОТЕ
Р
 ассмотрите на препарате «Общая морфология клетки» клетки печени при малом увеличении, а затем при большом. Зарисуйте при большом увеличении пять-шесть печеночных клеток, обозначив на рисунке цитоплазму, ядро и границы клеток (цитоплазма - розового цвета, ядро - фиолетового, одно-два темно-фиолетовых ядрышка).
ассмотрите на препарате «Общая морфология клетки» клетки печени при малом увеличении, а затем при большом. Зарисуйте при большом увеличении пять-шесть печеночных клеток, обозначив на рисунке цитоплазму, ядро и границы клеток (цитоплазма - розового цвета, ядро - фиолетового, одно-два темно-фиолетовых ядрышка).
Рис 46 Общая морфология клетки 1 – ядра печеночных клеток; 2 – цитоплазма; 3 – пикнотические ядра
* Найдите на препарате пикнотически измененные ядра, сравните характер хроматина пикнотических ядер.
Н
 а препарате «Хондриосомы в клетках эпителия кишечника аскариды» найдите эпителиальные клетки. При большом увеличение рассмотрите эпителиоциты, обратите внимание на ярко выраженную полярность клеток. На апикальном полюсе клетки найдите микроворсинки, зону пиноцитозных пузырьков, зону митохондриального ретикулюма - хондриосом, которые находятся в виде розовой «пыли».
а препарате «Хондриосомы в клетках эпителия кишечника аскариды» найдите эпителиальные клетки. При большом увеличение рассмотрите эпителиоциты, обратите внимание на ярко выраженную полярность клеток. На апикальном полюсе клетки найдите микроворсинки, зону пиноцитозных пузырьков, зону митохондриального ретикулюма - хондриосом, которые находятся в виде розовой «пыли».
* Рис 47 Хондриосомы в клетках эпителия кишечника аскариды 1 – базальная мембрана; 2 – ядра; 3 - митохондрии (темные точки) 4 – пикнотические пузырьки; 5 – зона микроворсинок
В центральной части клетки обратите внимание на мелкие яркие пузырьки разного диаметра. Какие это органоиды? Какую функцию выполняют? Зарисуйте 3 - 4 клетки, обозначив на рисунке ядро, митохондрии, лизосомы, микроворсинки, базальную мембрану.

На препарате «Аппарат Гольджи в клетках спинального ганглия котенка» при большом увеличении найдите в нейроцитах расположенный вокруг ядра комплекс Гольджи.
* Рис.48 Аппарат Гольджи в клетках спинального ганглия котенка 1 – комплекс Гольджи; 2 – ядро; 3 - ядрышко
Определите тип аппарата Гольджи - диффузный или пластинчатый. Зарисуйте 2 - 3 клетки, обозначив ядро, ядрышко и комплекс Гольджи.
Н
 а препарате «Влючения жира в клетках печени аксолотля» найдите при малом увеличении уже знакомые клетки, в данном случае специально обработанные для выявления жировых включений (черные капли различной величины). Зарисуйте 5 - 6 клеток, обозначив ядра, границы клеток, жировые включения.
а препарате «Влючения жира в клетках печени аксолотля» найдите при малом увеличении уже знакомые клетки, в данном случае специально обработанные для выявления жировых включений (черные капли различной величины). Зарисуйте 5 - 6 клеток, обозначив ядра, границы клеток, жировые включения.
Рис. 49 Включения жира в клетках печени аксолотля 1 – жировые включения; 2 – ядра.
Р
 ассмотрите при малом увеличении препарат «Включения гликогена в клетках печени аксолотля». При большом увеличении найдите в клетках ядро и окрашенные кармином в красный цвет включения гликогена (в виде колпачков). Зарисуйте 5 - 6 клеток, обозначив ядро и включения гликогена.
ассмотрите при малом увеличении препарат «Включения гликогена в клетках печени аксолотля». При большом увеличении найдите в клетках ядро и окрашенные кармином в красный цвет включения гликогена (в виде колпачков). Зарисуйте 5 - 6 клеток, обозначив ядро и включения гликогена.
Рис. 50 Включения гликогена в клетках печени аксолотля 1 – пикнотические ядра; 2 – ядра печеночных клеток; 3 – цитоплазма печеночных клеток; 4 – включение гликогена
Н
Рис. 51 Амитоз в клетках мочевого пузыря мыши
1- ядра клеток
 а препарате «Амитоз в клетках мочевого пузыря мыши» при малом увеличении микроскопа найдите крупные с большими ядрами клетки. Внимательно рассмотрите ядро этих клеток при большом увеличении. Ядра различной формы. Одни - круглые, в других намечается перетяжка, в третьих перетяжка очень глубокая. Есть также клетки, где имеется два ядра и начинается раздвоение тела самой клетки. Зарисуйте клетки в различных стадиях амитотического деления.
а препарате «Амитоз в клетках мочевого пузыря мыши» при малом увеличении микроскопа найдите крупные с большими ядрами клетки. Внимательно рассмотрите ядро этих клеток при большом увеличении. Ядра различной формы. Одни - круглые, в других намечается перетяжка, в третьих перетяжка очень глубокая. Есть также клетки, где имеется два ядра и начинается раздвоение тела самой клетки. Зарисуйте клетки в различных стадиях амитотического деления. Н
 а препарате «Митоз в клетках корешка лука» найдите клетки на разных стадиях деления. Зарисуйте клетки на различных стадиях митоза.
а препарате «Митоз в клетках корешка лука» найдите клетки на разных стадиях деления. Зарисуйте клетки на различных стадиях митоза.
Рис. 52 Митоз в клетках корешка лука 1 – конец профазы; 2 - начало телофазы; 3 – анафаза; 4 – метафаза; 5 – завершение телофазы
практические задания и ситуативные задачи для проверки знаний и умений
В клетках хорошо развита гранулярная ЭПС и комплекс Гольджи. Какова основная функция этих клеток?
В клетках печени идет активный синтез гликогена и белков. Какие структуры должны быть наиболее развиты в этих клетках?
Нейтрофильный лейкоцит имеет псевдоподии (ложноножки) и большое количество лизосом. С каким процессом связано развитие этих структур?
При электронной микроскопии была обнаружена в клетках деструкция митохондрий. Какие процессы в клетках нарушены?
Ядро нервной клетки светлое с крупным интенсивно окрашенным ядрышком («глаз совы»). О чем это говорит?
Клетки каемчатого эпителия имеют множество выростов цитоплазматической мембраны верхней части клетки и большое количество митохондрий. Какой процесс осуществляют эти клетки? Что необходимо для данного процесса?
Рассмотрите два гистологических препарата «Митоз в корешках лука» и «Центросомы и ахроматиновое веретено деления в яйцеклетках лошадиной аскариды ». Найдите на обоих препаратах клетки в разных стадиях митоза, зарисуйте, отметив хромосомы, «метафазную пластинку», стадию «дочерних звезд». Отметьте отличительные особенности митоза в растительных и животных клетках.
Рассмотрите два препарата «Хондриосомы в эпителии кишечника аскариды» и «Мерцательный эпителий кишечника беззубки». Рассмотрите пласты эпителиальных клеток, обратите внимание на апикальный (обращенный наружу) полюс клеток, рассмотрите его с иммерсионным увеличением. Какие органеллы специального назначения можно дифференцировать в этих участках клеток? Опишите их морфофункциональные особенности.
Рассмотрите препарат «Тигроид в нервных клетках спинного мозга». На поперечном срезе спинного мозга видно серое вещество в виде бабочки, окруженное белым веществом. В передних рогах серого вещества находятся тела двигательных нейронов. В цитоплазме клеток хорошо различимы базофильные гранулы, которые были названы «тигроидом» - специальным образованием, присущим только нейронам. Что такое тигроид? Почему он проявляет базофилию (сродство к основным красителям)? Почему тигроида нет в других клетках? Длительное время тигроид считали «запасным питательным веществом», поскольку в различных нейронах он развит по-разному, и почти отсутствует в нервных клетках при патологии. Объясните, с чем связано различное развитие тигроида.
Рассмотрите препарат «Мезенхима зародыша курицы», найдите клетки зародышевой ткани. Отметьте морфологические особенности молодых быстрорастущих клеток. С чем связана базофилия цитоплазмы в этих клетках?
Дополнительная литература
Атлас сканирующей электронной микроскопии клеток, тканей и органов. /Под. ред. Волоковой, В.А. Шахламова, А.А. Миронова – М.: Медицина, 1987
О.В. Быков В.Л. Цитология и общая гистология (функциональная морфология клеток и тканей человека). – СПб.: СОТИС, 1998.
Джавец Э., Мельник ДЖ.Л., Эйдельберг Э.Руководство по медицинской микробиологии: – М.: Медицина, 1982
Леви А., Сикевиц Ф. Структура и функции клетки. – М.: Мир, 1971.
Международная гистологическая номенклатура / Под. ред. В.В.Семченко, Р.П. Самусева, М.В.Моисеева и З.Л. Колосовой. – Омск: Омская медицинская академия, 1999
Меркулов Г.А. Курс патологогистологической техники. – Изд-во Медицина, Ленинградское отделение, 1969.
Рейва П., Эверт Р., Айкхрн С. Современная ботаника: в 2-х т. Т.1.: - М., Мир, 1990
Ченцов Ю.С. Общая цитология. – М.: Изд-во МГУ, 1995
Ченцов Ю.С. Введение в клеточную биологию. – М.: ИКЦ «Академкнига», 2004
Хэм А., Кормак Д. Гистология: - М., Мир, 1982 –Т.1.
Шлегель Г. Общая микробиология:- М., Мир, 1987
Щелкунов С.И. Клеточная теория и учение о тканях. – Медгиз, Ленинградское отделение, 1958.
67
Просмотр содержимого документа
«клетка модуль 7»
Раздел 3. основы физиологии клетки
При изучении этого раздела Вы узнаете:
сущность понятий обмен веществ, ассимиляция, диссимиляция;
основные этапы обмена веществ;
сущность реакций энергетического обмена;
этапы и механизм процессов фотосинтеза;
сущность процессов биосинтеза белка;
механизм процессов транскрипции и трансляции;
этапы жизненного цикла клетки;
сущность процессов роста и размножения клеток;
сущность процессов митоза и его стадий.
ИНФОРМАЦИОННЫЙ БЛОК
3.1. Клеточный метаболизм.
Метаболизм, или обмен веществ - характерный признак всех живых существ. Без обмена веществ жизнь невозможна. В организме, его органах, тканях, клетках непрерывно образуются, разрушаются и обновляются их структурные компоненты. Для образования новых клеток, для работы всех органов и систем органов, а также для совершения физической работы организму нужна энергия. Эта энергия образуется в процессе обмена веществ. Источником энергии служат питательные вещества, поступающие с пищей, а у фототрофных организмов источником энергии является свет - энергия Солнца.
Непрерывность обмена веществ и энергии возможна потому, что организм существует как открытая система. Это значит, что с пищей, водой, при газообмене из внешней среды в организм поступают различные соединения. По химическому составу они отличаются от внутренней среды данного организма. В организме эти соединения подвергаются изменениям и превращениям, и, в конце концов, становятся тождественными химическим веществам организма и входят в его морфологические структуры. Через определенный период усвоенные вещества подвергаются разрушению, выделяя скрытую в них энергию, а конечные продукты расщепления выводятся во внешнюю среду. При этом разрушенную молекулу сразу же замещает новая, не нарушая структурной целостности организма. Следовательно, организм - не статичная, а открытая поточная система.
Поток вещества и энергии, наблюдаемый в организме, обусловливает самообновление и самовоспроизведение, которые осуществляются на всех уровнях организации - от молекулярного до организменного.
Обмен веществ складывается из двух неразрывных и взаимосвязанных процессов - ассимиляции (или анаболизма) и диссимиляции (или катаболизма).
Ассимиляция или анаболизм - это реакции биологического синтеза сложных молекул органических соединений, специфичных для данного организма, из более простых компонентов, поступающих в клетки. Например, из аминокислот в анаболических процессах синтезируются пептиды или белки. Анаболизм является основой для построения новых клеточных структур, для формирования новых тканей в процессе роста и поскольку за счет данных реакций идет образование тканевых и клеточных структур – эти реакции называются также пластическим обменом.
Энергия для анаболических реакций образуется в реакциях диссимиляции или катаболизма, при которых происходит расщепление молекул сложных органических соединений с выделением энергии. К реакциям катаболизма относят расщепление белков до аминокислот, жиров до жирных кислот и глицерина, сахаров до глюкозы, а также и реакции расщепления мономеров до неорганических соединений (углекислого газа, мочевины, воды и др.) Большинство перечисленных реакций идет с выделением энергии, заключенной в химических связях, поэтому эти реакции составляют часть энергетического обмена. Конечные продукты катаболизма не могут быть окислены в организме и удаляются из него.
Соотношение процессов катаболизма и анаболизма определяет три различных состояния: динамическое равновесие, рост и частичное разрушение структур организма. При динамическом равновесии анаболизм и катаболизм уравновешены. Это норма для взрослого зрелого и здорового организма. Преобладание анаболизма ведет к росту, это состояние является нормой для детей. Преобладание катаболизма приводит к уменьшению массы, истощению организма. Это лежит в основе старения или характерно для болезни.
Обмен веществ состоит из трех этапов: подготовительного, клеточного и этапа выделения.
Клеточный этап начинается с поступления в клетку из тканевой жидкости мономеров: аминокислот, глюкозы, жирных кислот и глицерина и др. Эти вещества в клетку поступают путем активного транспорта - с помощью ферментов клеточной мембраны (см. раздел 2). В клетке из аминокислот образуется необходимый клетке белок. Этот процесс происходит на рибосомах (как свободных, так и связанных с ЭПС). Синтезированные белки клетки являются или структурными - из них строятся клеточные органеллы, или ферментами, которые обеспечивают все процессы жизнедеятельности, а также выполняют другие функции. Часть аминокислот подвергается расщеплению, конечными продуктами которого являются CO2, H2O NH3. Из мономеров на каналах гладкой ЭПС синтезируются собственные жиры (фосфолипиды, стероиды и др.) и углеводы. В ядре идут постоянно процессы репликации и транскрипции - синтеза ДНК и РНК.
Для всех процессов жизнедеятельности необходима энергия, которая образуется в процессе реакций катаболизма.
Процессы ассимиляции и диссимиляции связаны не только структурно, т.е. осуществляются в клеточных органеллах, но и биохимически: продукты реакций ассимиляции могут быть использованы клеткой для энергетического обмена. Например, если в клетку не поступают углеводы и жиры для энергетического обмена, клетка расщепляет структурные белки.
3.2. Ферментативный характер реакций обмена
Большую роль в протекании реакций внутриклеточного обмена играют многочисленные клеточные ферменты. Ферменты - это высокоэффективные биологические катализаторы, белковые молекулы, синтезируемые живыми клетками. В каждой клетки имеются сотни и тысячи различных ферментов. С их помощью осуществляются разнообразные химические реакции, лежащие в основе процессов жизнедеятельности.
Ферменты обычно состоят из нескольких полипептидных цепей и обладают четвертичной структурой. Кроме того, ферменты могут иметь в своем составе и небелковые компоненты. Белковая часть называется апоферментом, а небелковая - кофактором (если это простое неорганическое вещество) или коферментом (если это сложное органическое соединение). Предшественниками многих коферментов являются витамины. Апофермент и кофермент составляют единый каталитически активный комплекс - голофермент.
Биологические катализаторы имеют несколько общих свойств:
по структуре они - глобулярные белки;
увеличивают скорость реакции, но сами в этой реакции не расходуются;
присутствие фермента никак не влияет на конечный продукт реакции;
очень малая концентрация фермента оказывает специфическое действие в отношении большого количества субстрата за малый промежуток времени;
ферменты обладают специфичностью.
Виды специфичности: абсолютная - один фермент катализирует превращения только одного субстрата; относительная - один фермент катализирует превращения нескольких, сходных по химическому строению, веществ; специфичность пути превращения - фермент катализирует только определенный путь превращения своего субстрата, то есть определенную химическую реакции; и стереоспецифичность - фермент обеспечивает превращения одного из стереоизомеров (например, или цис-, или транс-изомера).
Активность ферментов и скорость ферментативной реакции зависит от многих факторов.
Во-первых, от концентрации фермента - при возрастании концентрации фермента скорость реакции возрастает прямо пропорционально увеличению концентрации фермента. Во вторых, от концентрации субстрата - при ее увеличении сначала скорость возрастает прямо пропорционально увеличению концентрации, а затем этот процесс замедляется, и при достижении определенной концентрации субстрата скорость реакции больше не возрастает и остается постоянной.
Активность фермента зависит также от температуры - при повышении температуры на каждые 10° скорость возрастает примерно в 2 раза, но до определенного предела. Температура, обеспечивающая наибольшую скорость реакции и активность фермента, называется оптимальной. При дальнейшем повышении температуры до 39-50°С практически все ферменты денатурируют и теряют свойства. Ферментативная активность зависит и от реакции среды (pH). То значение pH, при котором скорость химической реакции максимальна, считается оптимальным для данного фермента. Например, для ферментов крови оптимальное значение pH=7,36.
В контакт с субстратом вступает лишь небольшая часть молекулы фермента, называемая активным центром. Форма активного центра фермента динамически соответствует субстрату, что можно сравнить с перчаткой, меняющей свою форму при надевании на руку. Активный центр состоит из двух участков - контактного и каталитического. Первый отвечает за связывание фермента с молекулой субстрата, а второй за протекание собственно катализа.
Известны различные соединения, которые могут тормозить химические реакции. Такие соединения называются ингибиторами. Ингибирование может быть обратимым и необратимым. В первом случае ингибитор непрочно связан с ферментом и может отделяться. Во втором случае ингибитор прочно связывается с ферментом, активность которого полностью утрачивается.
3.3. Фотосинтез
Жизнедеятельность всех организмов должна обеспечиваться энергией. Энергия эта поступает из окружающей среды. Первичным источником энергии для всего живого служит солнечный свет. Эта энергия улавливается растениями в процессе фотосинтеза и превращается в химическую энергию, которая сохраняется в запасных веществах, образующихся при фотосинтезе.
Впервые решить проблему питания растений попытался Я. Б. ван Гельмот более 300 лет назад. Он выращивал дерево в горшке, и только поливая его, добился значительного увеличения массы, однако масса почвы уменьшилась лишь на несколько грамм. Д. Пристли 17 августа 1771 года провел целую серию опытов по «улучшению» воздуха растениями. Детально изучил процессы фотосинтеза К. А.Тимирязев. В своей работе «Солнце, жизнь и хлорофилл» он не только описал механизм реакций фотосинтеза, но и показал значение зеленых растений для жизни на Земле.
Фотосинтез осуществляется различными группами автотрофных организмов - высшими зелеными растениями, многоклеточными зелеными, бурыми и красными водорослями, некоторыми одноклеточными эукариотами (например, эвгленой зеленой, хламидомонадой и другими) и бактериями. Фотосинтез - это единственный процесс в природе, который идет с запасанием химической энергии и обеспечивает доступным источником ее все существующие в природе организмы. Биологическая роль фотосинтеза огромна. В течение года в результате фотосинтеза на Земле усваивается около 200 млрд. тонн углекислого газа, образуется около 100 млрд. тонн органического вещества и выделяется около 145 млрд. тонн молекулярного кислорода. Фотосинтез, осуществляемый первыми фотосинтезирующими организмами (сине-зелеными водорослями), вызвал появление свободного кислорода в атмосфере и создал озоновый экран, обеспечив, таким образом, условия для биологической эволюции.
У всех фотосинтезирующих организмов, за исключением бактерий, процесс фотосинтеза представляет собой образование органических веществ из углекислого газа и воды с использованием энергии солнечного света, сопровождающийся выделением кислорода.
С hν
nH2O + nCO2 → (CH2O)n + nO2,
где hν - квант света, и если n = 6, то это соответствует образованию в качестве продукта фотосинтеза молекулы моносахарида глюкозы - C6H12O6.
Фотосинтезирующие бактерии также синтезируют на свету органические вещества из углекислого газа, однако, вместо воды они используют другие доноры водорода - неорганические (например, сероводород) или органические (например, молочная кислота) соединения.
Для фотосинтезирующих бактерий, донором водорода у которых служит сероводород (например, зеленые серные бактерии), общее уравнение фотосинтеза выглядит следующим образом:
hν
2nH2S + nСO2 → (CH2O)n + nH2O + 2nS,
где (CH2O) - фрагмент молекулы углевода.
Во всех случаях при фотосинтезе преобразование световой энергии в химическую включает окислительно-восстановительный перенос электронов и сопряженное с ним образование АТФ.
Фотосинтетические процессы у высших растений и некоторых водорослей осуществляется в особых клеточных органеллах - хлоропластах, строение которых рассматривалось в разделе 2.
Фотосинтезирующие бактерии содержат другие специализированные образования - хроматофоры, в мембранах которых также локализованы пигменты и специфические ферменты для реакций фотосинтеза.
Фотосинтез включает в себя три основных принципиально различных типа химических реакций: фотофизические, фотохимические и ферментативные. Первые два типа реакций входят в так называемую световую фазу фотосинтеза, для осуществления которой необходим свет, а последние реакции составляют темновую фазу фотосинтеза, свет для которой не нужен.
Реакции световой фазы фотосинтеза протекают на внутренних мембранах хлоропластов. Фотосинтетические пигменты, главный из которых хлорофилл, особым образом сгруппированы на этих мембранах, и формируют так называемые фотосистемы. Расположение молекул пигментов в фотосистеме напоминает пирамиду.
Ф отон света может улавливать любая из молекул. При этом один из подвижных электронов этой молекулы хлорофилла переходит в возбужденное состояние, а затем отделяется и переходит к другой молекуле фотосистемы. Заканчиваются процессы перемещения электрона тем, что он оказывается у молекулы, расположенной на вершине пирамиды - в хлорофилле-ловушке.
отон света может улавливать любая из молекул. При этом один из подвижных электронов этой молекулы хлорофилла переходит в возбужденное состояние, а затем отделяется и переходит к другой молекуле фотосистемы. Заканчиваются процессы перемещения электрона тем, что он оказывается у молекулы, расположенной на вершине пирамиды - в хлорофилле-ловушке.
Если экстрагировать хлорофилл из растения и осветить его раствор, то раствор начнет флуоресцировать - поглощенная энергия будет высвечиваться при возвращении электронов в «основное состояние». В хлоропластах этого не происходит. Молекулы хлорофилла организованы в них таким образом, что возбужденные электроны не возвращаются сразу же в «основное состояние», а передаются другим молекулам - электрон отделяется от фотосистемы и движется по электронно-транспортной цепи. В ходе этого движения синтезируются АТФ и НАДФH.
Световая фаза идет при участии двух фотосистем.
В фотосистеме I при поглощении света хлорофиллом энергия электронов возрастает до уровня, на котором они могут восстанавливать неидентифицированный акцептор электронов X. От него электроны через ферредоксин (Fd) передаются НАДФ и восстанавливают его. Этот восстановленный НАДФ сам может затем поставлять электроны, необходимые для восстановления атмосферного СО2 до глюкозы. Таким путем световая энергия превращается в химическую. Источником протонов для синтеза НАДФH является вода.
В фотосистеме I имеет место также циклический поток электронов: электроны, захваченные акцептором X, возвращаются через цитохром h хлорофиллу. При таком потоке электронов синтезируется только АТФ.
В фотосистеме II электроны хлорофилла при поглощении света переходят на энергетический уровень, на котором они могут присоединяться к акцептору электронов Q-C550. Эти электроны передаются затем в серии окислительно-восстановительных реакций, включающих пластохинон, цитохром и пластоцианин, хлорофиллу фотосистемы I. В фотосистеме II богатые энергией электроны возбужденного хлорофилла передаются системе переноса электронов и замещаются бедными энергией электронами, поступающими от воды.
Параллельно процессу переноса электронов по электронно-транспортной цепи происходит процесс фотолиза воды:
hν
2 H2O…→ 2 [O] + 4 H+ + 4ē
О бразовавшиеся электроны замещают электроны, отданные молекулами хлорофилла, а атомы кислорода объединяются в молекулы и как побочный продукт удаляются в атмосферу.
бразовавшиеся электроны замещают электроны, отданные молекулами хлорофилла, а атомы кислорода объединяются в молекулы и как побочный продукт удаляются в атмосферу.
Хлорофилл, следовательно, играет роль промежуточного соединения на пути электронов от низкого энергетического уровня в молекуле воды к высокому энергетическому уровню в конечном акцепторе электронов. Именно в молекуле хлорофилла световая энергия используется для того, чтобы «перекачивать» электроны на этот высокий энергетический уровень.
Богатые энергией электроны из фотосистемы II, пройдя через ряд соединений, составляющих так называемую цепь переноса электронов, в конце концов, замещают электроны, утраченные хлорофиллом фотосистемы I. В цепи переноса электронов происходит ряд окислительно-восстановительных реакций, и в каждой из этих реакций электроны переходят на несколько более низкий энергетический уровень. Часть энергии, отдаваемой на отдельных этапах переноса электронов, используется для синтеза АТФ из АДФ и неорганического фосфата. Поэтому можно сказать, что синтез АТФ сопряжен с переносом электронов.
Еще одно место синтеза АТФ связано с фотосистемой I. Это циклическое фотофосфорилирование зависит от электронов, восстанавливающих акцептор X, а затем возвращаемых через ряд переносчиков хлорофиллу, от которого они первоначально и поступили.
Таким образом, в процессе фотосинтеза участвуют две фотосистемы, связанные между собой через систему переноса электронов и два богатых энергией соединения: АТФ и восстановленный НАДФ.
Эти соединения поставляют энергию для последующих реакций, не требующих участия света и приводящих, в конечном счете, к восстановлению СО2 до сахаров. Восстановление СО2 начинается с фиксации молекул СО2 молекулами «акцептора», после чего атомы углерода вступают в цикл реакций, в результате которого акцепторные молекулы регенерируют и на каждые шесть фиксированных молекул СО2 образуется одна молекула глюкозы. Именно в этих темновых реакциях фотосинтеза химическая энергия, которую генерируют световые реакции, стабилизируется в молекуле глюкозы. В конце концов, из молекул глюкозы образуется крахмал - соединение, в виде которого запасаются и атомы углерода, и энергия.
В ходе реакций темновой фазы происходят процессы фиксации углекислого газа при участии молекул НАДФH, на протекание которых затрачивается энергия АТФ. Эти реакции проходят во внутренней среде - строме хлоропластов. Наиболее распространенным способом фиксации углерода является цикл химических реакций, названный по имени его открывателя циклом Кальвина. Он включает в себя двеннадцать химических реакций. На каждый оборот цикла фиксируется одна молекула углекислого газа.
Суммарная реакция цикла Кальвина:
18АТФ + 12НАДФH + 6CO2 →C6H12O6 + 18H3PO4 + 18АДФ + 6H2O + 12НАДФ
Таким образом, суммарная реакция фотосинтеза имеет следующий вид:
6СО2 + 12Н2О + hν → С6 Н12О6 + 6О2 ↑ + 6Н2О
Невозможно переоценить значение фотосинтеза для всей жизни на Земле и для человечества в частности. Фотосинтез, в процессе которого улавливается и трансформируется энергия, не только поставляет «горючее», необходимое для существования всего живого; ему мы обязаны и теми энергетическими резервами, которыми располагает в настоящее время человечество с его все более усиливающейся индустриализацией, поскольку большая часть этих резервов представлена запасами ископаемого горючего. За счет фотосинтеза Земля получает ежегодно около 840 триллионов киловатт-часов, что более чем в 10 раз превышает мировое потребление энергии. Фотосинтез же поставляет и необходимый всем высшим формам жизни атмосферный кислород, который представляет собой его побочный продукт, высвобождающийся при окислении воды.
3.4. Энергетический обмен в клетке, его значение и этапы
Животные организмы, не способные к фотосинтезу, зависят от этих запасных веществ, синтезируемых растениями. Запасенная энергия переводится в форму, в которой она может использоваться растительными и животными клетками для выполнения какой-нибудь работы, например для синтеза других молекул, необходимых клетке, или же для механической, электрической и осмотической работы.
Эффективное высвобождение запасенной энергии происходит в процессе аэробного дыхания, во время которого молекулы, содержащие запасенную энергию, расщепляются в определенной последовательности реакций.
Все химические соединения характеризуются определенной энергией; часть энергии представляет собой так называемую свободную энергию, которая может быть использована для выполнения работы. Количество свободной энергии, которое может быть извлечено из данной молекулы, зависит от ее природы и от расположения атомов, входящих в ее состав. При химических реакциях, когда одни химические связи разрываются и вместо них возникают другие, происходит перераспределение энергии в молекуле. При некоторых реакциях свободная энергия высвобождается и может расходоваться на выполнение какой-нибудь работы. Существуют и реакции, идущие с потреблением свободной энергии.
Источником энергии в клетке чаще всего служат окислительно-восстановительные реакции, при которых происходит перенос электронов от одного соединения (которое, следовательно, окисляется) к другому (восстанавливающемуся). Способность отдавать электроны (т. е. действовать как восстановитель) определяет величину химического потенциала данного соединения. Когда соединение с более высоким химическим потенциалом восстанавливает соединение с более низким потенциалом, свободная энергия высвобождается. Разность между начальной и конечной энергиями электрона (или электронов) характеризует количество свободной энергии, «высвободившейся» в данной реакции. В клетке, разумеется, энергия в действительности не высвобождается; просто часть ее запасается в форме, которая затем может быть использована.
В переносе энергии от энерговыделеяющих химических реакций к процессам, идущим с потреблением энергии (которые собственно и составляют работу клетки), принимает участие аденозинтрифосфат, выполняющий важную роль носителя энергии. АТФ образуется в клетке путем присоединения еще одной фосфатной группы к аденозиндифосфату (АДФ). Поскольку э нергетический уровень АТФ выше, чем у АДФ, на присоединение этой фосфатной группы должна затрачиваться энергия. Именно та энергия, которая поставляется реакциями окисления-восстановления, затрачивается на образование АТФ и, следовательно, запасается в его молекуле. Это достигается благодаря сопряжению реакции фосфорилирования, т. е. переноса фосфатной группы, с реакциями окисления-восстановления (с переносом электронов).
нергетический уровень АТФ выше, чем у АДФ, на присоединение этой фосфатной группы должна затрачиваться энергия. Именно та энергия, которая поставляется реакциями окисления-восстановления, затрачивается на образование АТФ и, следовательно, запасается в его молекуле. Это достигается благодаря сопряжению реакции фосфорилирования, т. е. переноса фосфатной группы, с реакциями окисления-восстановления (с переносом электронов).
Энергия, запасенная в молекуле АТФ, высвобождается, когда она требуется для выполнения какой-нибудь работы, в результате гидролиза АТФ до АДФ и неорганического фосфата. Для того чтобы эта свободная энергия не рассеивалась полностью в виде тепла, превращение АТФ в АДФ, совершающееся в клетке ферментативным путем, происходит таким образом, что часть энергии используется для выполнения работы. АТФ действует как своего рода энергетический «челнок». Реакции окисления-восстановления и сопряженное с ними фосфорилирование протекают при дыхании (когда запасенная в клетке энергия используется на образование АТФ) и при фотосинтезе (когда энергия, уловленная хлоропластами, поступает - через АТФ - в конечные продукты фотосинтеза, т.е. в запасные вещества растений).
Основным энергетическим веществом клетки является глюкоза. Процесс ее расщепления в клетке до воды и углекислого газа в присутствии свободного кислорода состоит из трех этапов:
Гликолиз и окислительное декарбоксилирование пировиноградной кислоты с образованием ацетил-КоА.
Окисление ацетил-КоА в цикле трикарбоновых кислот (цикле Кребса).
Окислительное фосфорилирование в электронно-транспортной цепи.
На первом этапе происходит образование двух молекул ацетилкоэнзима А (ацетил-КоА) из молекулы глюкозы. Это происходит в две стадии. На первой - молекула глюкозы расщепляется до двух молекул пировиноградной кислоты (ПВК). Эта стадия известна под названием гликолиза (гр glycos - сладкий lysis - расщепление). Он протекает в цитоплазме и не требует присутствия кислорода. Гликолиз состоит из одиннадцати последовательных реакций и осуществляется при помощи десяти ферментов.
Суммарное уравнение гликолиза:
C 6H12O6 + 2H3PO4 + 2АДФ + 2НАД+ → 2CH3COCOOH + 2АТФ + 2НАДH
6H12O6 + 2H3PO4 + 2АДФ + 2НАД+ → 2CH3COCOOH + 2АТФ + 2НАДH
Дальнейшая судьба ПВК зависит от наличия в среде кислорода. В бескислородных условиях образовавшуюся пировиноградную кислоту анаэробные организмы перерабатывают либо в молочную кислоту (молочнокислое брожение), либо в этиловый спирт (спиртовое брожение).
У аэробных организмов ПВК переносится из цитоплазмы в митохондрии и там начинается вторая стадия первого этапа - окислительное декарбоксилирование ПВК. В этих реакциях участвуют три фермента и пять коферментов.
Уравнение реакции окислительного декарбоксилирования пировиноградной кислоты с образованием ацетил-КоА:
CH3COCOOH + КоА + НАД+ → CH3CO-КоА + CO2 + НАДH
Ацетил-КоА поступает в цикл трикарбоновых кислот, где происходит полное окисление промежуточных продуктов. Именно в матриксе митохондрии располагаются ферменты цикла Кребса. В целом цикл Кребса состоит более чем из десяти реакций.
Суммарное уравнение реакций цикла трикарбоновых кислот:
CH3CO-КоА + АДФ + Ф + 3 НАД+ + ФАД+ → КоА + АТФ + 3НАДH + ФАДH2 + 2CO2
В
осстановленные НАД и ФАД участвуют в третьем этапе клеточного дыхания - окислительном фосфорилировании.
В
ходе третьего этапа происходит окисление атомов водорода молекулярным кислородом с образованием воды и запасанием выделившейся энергии в виде АТФ. Процесс окислительного фосфорилирования осуществляется в несколько стадий путем последовательной передачи электронов по цепи молекул переносчиков или электронно-транспортной цепи. Эти молекулы локализуются на внутренней мембране митохондрий.
В результате работы электронно-транспортной цепи атомы водорода с НАДH и ФАДH2 разделяются на протоны и электроны. Электроны в конечном итоге переносятся на атомы кислорода, а протоны - в межмембранное пространство митохондрий. Затем протоны через пронизывающий всю мембрану комплекс фермента АТФ-синтетазы возвращаются в матрикс митохондрий. При этом образуется АТФ из АДФ и фосфорной кислоты. Механизм образования АТФ объясняется наиболее распространенной хемоосмотической теорией Митчелла, согласно которой АТФ образуется за счет кинетической энергии протона, проходящего через АТФ-синтетазу.
Суммарное уравнение всех реакций расщепления глюкозы будет выглядеть следующим образом:
C6H12O6 + 6O2 + 4 АДФ + 4 Ф + 10 НАД+ + 2 ФАД+→
→ 6 CO2 + 6 H2O + 4 АТФ + 10 НАДH + 2 ФАДH
Одна молекула НАДH2 способна образовать три молекулы АТФ, а одна молекула ФАДH2 - две молекулы АТФ. Следовательно, суммарное уравнение расщепления глюкозы можно записать следующим образом:
C6H12O6 + 6 O2 + 38 H3PO4 + 38 АДФ → 6 CO2 + 44 H2O + 38 АТФ
АТФ транспортируется ко всем участкам клетки, где необходима энергия. Эта энергия, заключенная в макроэргических связях в молекуле АТФ, выделяется при расщеплении АТФ до АДФ. АДФ снова поступает в митохондрии, где в ходе восстановительных реакций превращается в АТФ, связав энергию, освобожденную при окислении веществ.
Углекислый газ удаляется у растений через устьица, а у животных - через органы дыхания.
Кроме глюкозы окислению могут быть подвергнуты моносахариды, все 20 аминокислот, а также жирные кислоты и спирты липидов. Кроме указанного, существует еще несколько путей окисления органических веществ, например, пентозофосфатный цикл, в котором образуется НАДФH2, широко используемый в процессах биосинтеза.
3.5. Пластический обмен
Пластическим обменом называют все реакции биосинтеза, идущие в клетках организма. Это синтез белков, жиров, углеводов, нуклеиновых кислот. За счет процессов ассимиляции происходит образование клеточных структур, рост клеток, а, следовательно, всего организма. Синтетические процессы происходят на мембранах эндоплазматической сети и комплекса Гольджи. На гранулярной ЭПС идет синтез белка и фосфолипидов мембран, агранулярной ЭПС - синтез углеводов и липидов. Синтез нуклеиновых кислот идет в ядре и носит матричный характер. Процессы репликации (удвоения ДНК) катализируются ДНК-полимеразой, иРНК транскрибируется на ДНК, синтез рРНК идет в ядрышке.
3.5.1. Синтез белка
В настоящее время известно, что сходство и различие организмов определяется главным образом набором белков. Чем ближе организмы по происхождению друг к другу, тем большее сходство белков они обнаруживают. Молекула ДНК контролирует деятельность клетки и обеспечивает передачу наследственной информации в поколениях. Синтез белка происходит в рибосомах, расположенных в цитоплазме, а молекула ДНК находится главным образом в ядре клетки, таким образом, молекулы ДНК непосредственного участия в синтезе белка не принимают. В ДНК содержится и хранится информация о структуре белков, причем порядок расположения нуклеотидов в ДНК определяет порядок аминокислот в молекуле белка, то есть его первичную структуру. Информация, которую содержит молекула ДНК, называется генетической. Исследования показали, что четыре вида нуклеотидов и порядок их сочетания в молекуле ДНК представляют собой код, который определяет биосинтез специфических белков.
Код - это система символов, знаков, необходимая для перевода одной формы информации в другую. Каждую аминокислоту в молекуле белка определяет последовательность трех нуклеотидов ДНК, получивших название триплета или кодона. Участок ДНК, ответственный за синтез молекулы одного белка, называется геном. В состав гена входит столько триплетов, сколько аминокислот содержится в данном белке.
Код нуклеиновых кислот имеет свои особенности. Во-первых, триплетность - одну аминокислоту кодируют три нуклеотида. Это необходимо для обеспечения возможности определить все аминокислоты (четыре нуклеотида могут образовывать 64 разные тройки). Некоторые аминокислоты кодируются несколькими триплетами. Существуют бессмысленные кодоны (У-А-А; У-А-Г; У-Г-А), определяющие конец полипептидной цепи (стоп-кодоны).
Второй особенностью генетического кода является однозначность - код не перекрывается и один триплет кодирует только одну аминокислоту. Вырожденность генетического кода определяет способность кодировать одну и ту же аминокислоту (кроме метионина и триптофана) одновременно несколькими триплетами (от 2 до 6), что повышает устойчивость к случайным ошибкам (точечным мутациям).
Универсальность генетического кода - это способность триплетов кодировать одни и те же аминокислоты у самых разных организмов от вирусов до человека. Это является еще одним доказательством единства происхождения всего живого.
3.5.1.1 Этапы биосинтеза белка.
Биосинтез белка состоит из двух процессов: транскрипции и трансляции.
Транскрипция - синтез иРНК на ДНК, идет по принципу комплиментарности с помощью фермента РНК-полимеразы. Обычно копируется один или несколько рядом стоящих генов (ферментов, катализирующих один каскад реакций) - этот участок называется опероном. Опероны выявлены только у бактерий. В начале оперона имеется «посадочная площадка» для фермента РНК- полимеразы - эта площадка называется промотор. В транскрипции различают четыре стадии: связывание, инициация синтеза, элонгация синтеза и терминация.
Информация снимается с одной цепи.
Образовавшаяся цепочка иРНК подвергается процессу сплайсинга: отдельные некодирующие участки интроны вырезаются, а оставшиеся (кодирующие) - экзоны остаются и сшиваются. Зрелая иРНК поступает через ядерные поры в цитоплазму.
Т рансляция - перевод информации с кода триплетов иРНК в последовательность аминокислот полипептидной цепи. Трансляция идет на рибосомах с помощью транспортной РНК (тРНК), которая содержит 60-100 нуклеотидов. Несколько комплиментарных участков цепи тРНК слипаются за счет водородных связей и образуют крестообразную структуру. К одному концу тРНК прикрепляется аминокислота, противоположный конец - антикодон определяет аминокислоту, с которой соединяется акцепторный участок.
рансляция - перевод информации с кода триплетов иРНК в последовательность аминокислот полипептидной цепи. Трансляция идет на рибосомах с помощью транспортной РНК (тРНК), которая содержит 60-100 нуклеотидов. Несколько комплиментарных участков цепи тРНК слипаются за счет водородных связей и образуют крестообразную структуру. К одному концу тРНК прикрепляется аминокислота, противоположный конец - антикодон определяет аминокислоту, с которой соединяется акцепторный участок.
Т Рис. 60. Общая схема биосинтеза белка
В
ходе элонгации рибосома сидит на участке кодирующей последовательности и-РНК и удерживает молекулу пептидил-тРНК. Пептидил-тРНК - это растущая полипептидная цепь, ковалентно присоединенная к тРНК, которая и принесла последний аминокислотный остаток растущему пептиду. Когда пептидил-тРНК занимает Р-участок рибосомы (часть рибосомы, связывающая пептидил-тРНК), рибосома может связывать молекулу аминоацил-тРНК, соответствующую кодону, установленному на данный момент в А-участке (участок рибосомы связывающий аминоацил-тРНК). В результате, удерживаемая рибосомой пептидил-тРНК и вновь связанная аминоацил-тРНК оказываются в рибосоме бок о бок. Рибосома катализирует реакцию между этими двумя субстратами рибосомы - пептидил-тРНК и аминоацил-тРНК. Пептидильный остаток переносится от «своей» тРНК на аминогруппу аминоацил-тРНК, тем самым, удлиняясь на одну аминокислоту на С-конце. Теперь в Р-участке осталась деацилированная тРНК (без аминокислоты), а в А-участке помещается остаток тРНК удлиненной пептидил-тРНК. Следующий за этим акт транслокации состоит в том, что деацилированная тРНК (без аминокислоты) выталкивается из Р-участка, а пептидил-тРНК (ее остаток тРНК) перемещается вместе со связанным с ней кодоном иРНК из А-участка в Р-участок. В итоге А-участок освобождается, и в нем устанавливается следующий кодон иРНК. Цикл завершился.
Повторение таких циклов по числу кодонов мРНК создает полный процесс элонгации.
Достигнув специального нуклеотидного триплета (стоп-кодона, или кодона терминации), рибосома освобождает синтезированную полипептидную цепочку белка, происходит терминация трансляции.
Рибосомные частицы, закончившие трансляцию и отделившиеся от иРНК, снова готовы инициировать трансляцию.
П
Рис. 62 Схема цикла трансляции
осле инициации, двигаясь по цепи иРНК, рибосома все больше отодвигается от 5'-конца, и в результате ранее занимаемый ею отрезок иРНК становится свободным и способным связывать другую рибосому. Эта вторая рибосома тоже начинает трансляцию и двигается вслед за первой рибосомой, отодвигается от 5'-концевого отрезка и дает возможность третьей рибосоме сесть на иРНК и начать трансляцию и т.д. Таким путем, двигаясь вдоль иРНК друг за другом, несколько рибосом одновременно считывают одну и ту же запись и, следовательно, синтезируют идентичные полипептидные цепи. Такая структура, в которой одна иРНК связана со многими рибосомами, ее одновременно транслирующими, называется полирибосомой
Никакого отдельного от рибосомы белка-фермента, катализирующего образование пептидных связей на рибосоме, не существует. Итак, в процессе биосинтеза белка рибосома:
принимает кодированную генетическую информацию от ДНК в виде иРНК и расшифровывает ее;
катализирует образование пептидных связей в реакции транспептидации;
передвигает цепь иРНК и молекулы тРНК.
Таким образом, рибосома - сложная белоксинтезирующая частица, обладающая одновременно генетической, ферментативной и механической функциями.
3.5.1.2. Регуляция синтеза белка
Клетки разных тканей одного организма отличаются разным набором ферментов и других белков. Но все эти разные клетки произошли из одной оплодотворенной яйцеклетки в результате множества делений. Перед каждым в клетке происходит процесс удвоения ДНК - репликация. Следовательно, во всех клетках тела имеется одинаковый набор молекул ДНК - одна и та же генетическая информация о составе и структуре белков. Однако в каждой клетке реализуется не вся а только часть генетической информации. Механизмы регуляции синтеза отдельных белков изучили в 1961 году Франсуа Жакоб, Андре-Мишель Львов, Жан Люсьен Моно у бактерий. За эту работу они получили Нобелевскую премию.
У прокариот можно выделить структуру, называемую опероном. Это участок ДНК, состоящий из следующих частей:
П

Рис. 63. Регуляция биосинтеза белка у прокариот
А- образование открытого комплекса ДНК с РНК-полимеразой, начало транскрипции;
Б – одновременный процесс транскрипции и трансляции, образование полирибосом;-
В – индуктор связывается с белком-репрессором, который на может тормозить синтез иРНК
Г – молекула ДНК изгибается, белок- активатор вступает в контакт с РНК-полимеразой, активизируя ее.
ромотор - определенная последовательность нуклеотидов, которая связывается с ферментом РНК-полимеразой. Процесс связывания является первым этапом в последующей транскрипции.Оператор - участок ДНК, связанный с белком-репрессором. До тех пор, пока эта связь не нарушена, РНК-полимераза не будет считывать информацию.
Ряд структурных генов, кодирующих информацию о ферментах, участвующих в расщеплении определенного субстрата.
При поступлении субстрата в клетку его молекулы связываются с белком-репрессором, который после этого теряет способность взаимодействовать с оператором, происходит инициация, а РНК-полимераза начинает синтезировать иРНК, комплементарную структурным генам (элонгация). Затем в рибосомах синтезируются соответствующие ферменты, расщепляющие данный субстрат. Если субстрата не остается, освобождается белок-репрессор, который вновь блокирует оператор, и синтез иРНК и ферментов прекращается. Таким образом, бактерия синтезирует только те ферменты, которые ей необходимы в данный момент, что позволяет экономить энергию.
У эукариот регуляция активности генов значительно сложнее. На нее влияют гормоны, медиаторы, другие биологически активные вещества, причем транскрипция, выход иРНК и трансляция регулируются отдельно и могут быть разделены во времени.
85
Просмотр содержимого документа
«клетка модуль 8»
3.6. Деление клеток Митоз
Одно из важнейших положений клеточной теории гласит, что клетки не возникают de novo, а образуются путем размножения предшествующих клеток. Совершенствование гистологических методов и микроскопической техники позволило выявить важную роль ядра и в особенности заключенных в нем хромосом в процессах деления клеток. В 1879 году Бовери и Флемминг описали происходящие в ядре события, в результате которых образуются две идентичные клетки, они этот процесс назвали кариокинезом (carion - ядро, орех, cinesis - движение). В 1887 году Вейсман высказал мысль о том, что при образовании гамет происходит деление иного типа. В настоящее эти два типа называют соответственно митозом и мейозом. Период существования клетки от момента ее образования путем деления материнской клетки до собственного деления или смерти называется клеточным циклом. В жизненный цикл входит период выполнения клеткой многоклеточного организма специализированных функций, а также периоды покоя. В периоды покоя судьба клетки неопределенна: она может либо начать подготовку к митозу, либо встать на путь специализации. Значительная специализация делает невозможным деление клетки, и она после выполнения специализированных функций погибает.
Клетки, сохранившие способность к митозу, имеют клеточный цикл, ограниченный последовательностью деления. Период между следующими друг за другом делениями в таких клетках называют интерфазой. В интерфазе выделяют следующие периоды:
Пресинтетический период (G1). Клетка имеет полный диплоидный набор хромосом: 2n2c (гаплоидный - nc). В этот период происходят процессы биосинтеза белка, трансформации энергии и ее накопления в виде макроэргических соединений. Хромосомы в ядре деспирализованы. Клетка растет, увеличивается в размерах.
Синтетический период (S). Клетка готовится к предстоящему митозу. Происходит процесс репликации ДНК по принципу комплиментарности, то есть начинается процесс передачи наследственной информации. Генетическая характеристика клетки в конце синтетического периода 2n4c.
Постсинтетический период (G2). В этот период клетка морфологически содержит 2 молекулы ДНК. Генетическая характеристика клетки 2n4c. На этой стадии происходит накопление высокоэнергетических соединений, синтез белка, образование дочерней центриоли.
3.6.1. Митоз
Митоз (кариокинез, непрямое деление клетки) - наиболее распространенный способ репродукции клеток. Митоз обеспечивает возможность образования генетически равноценных клеток и сохраняет преемственность хромосом в ряду клеточных поколений.
Общая морфология митотических хромосом
Х ромосомы всех эукариотических клеток построены по одному плану. Они включают в себя: собственно тело - плечо, метомерный или конечный участок, и центромеру. Хромосомы представляют собой палочковидные структуры разной длины с довольно постоянной толщиной, обычно имеют два хромосомных плеча, соединенных в зоне центромеры. Эта зона называется первичной перетяжкой. Оба плеча хромосомы оканчиваются теломерами. Хромосомы с равными или почти равными плечами называют метацентрическими, с плечами неодинаковой длины - субметацентрическими. Палочковидные хромосомы с очень коротким плечом - акроцентрические.
ромосомы всех эукариотических клеток построены по одному плану. Они включают в себя: собственно тело - плечо, метомерный или конечный участок, и центромеру. Хромосомы представляют собой палочковидные структуры разной длины с довольно постоянной толщиной, обычно имеют два хромосомных плеча, соединенных в зоне центромеры. Эта зона называется первичной перетяжкой. Оба плеча хромосомы оканчиваются теломерами. Хромосомы с равными или почти равными плечами называют метацентрическими, с плечами неодинаковой длины - субметацентрическими. Палочковидные хромосомы с очень коротким плечом - акроцентрические.
В области первичной перетяжки (центромеры) расположен кинетохор - пластинчатая структура, имеющая форму диска. К нему подходят пучки микротрубочек митотического веретена. Эти пучки микротрубочек принимают участие в движении хромосом к полюсам клетки при митозе.
О Рис.64. Метафазная хромосома 1 – сестринские хромосомы (хроматиды); 2 – плечи хромосом; 3 – центромера; 4 – вто-ричная перетяжка – яд-рышковый организатор; 5 – теломеры; 6 – пучки микротрубочек.
В зоне первичной перетяжки присутствует особая, центромерная ДНК, отличающаяся высоким уровнем повторенности нуклеотидных последовательностей.
Некоторые хромосомы имеют вторичную перетяжку, которая обычно расположена вблизи дистального конца хромосомы и отделяет маленький участок - спутник. Вторичные перетяжки называют ядрышковыми организаторами, так как именно на этих участках в интерфазе происходит образование ядрышка. Теломерные концы хромосом не способны соединяться с другими хромосомами. В теломерах локализована особая теломерная ДНК, защищающая хромосому от укорачивания в процессе синтеза ДНК.
Выделяют четыре фазы митоза: профаза, метафаза, анафаза и телофаза.
Профаза - обычно самая продолжительная фаза клеточного деления. К началу профазы клетка обычно округляется, вязкость ее цитоплазмы нарастает. В раннюю профазу происходит скручивание, конденсация хроматина и образование хромосом. На хромосомах появляются перетяжки - центромеры - к ним в последствии прикрепляются нити веретена деления. Каждая хромосома состоит из двух свернутых спиральных нитей - хроматид, тесно прилегающих друг к другу. В раннюю профазу также исчезает ядрышко. В позднюю профазу центриоли расходятся по полюса клетки, а ядерная оболочка фрагментируется.
Генетическая характеристика 2n4c.
М етафаза. Хромосомы располагаются в экваториальной плоскости клетки (стадия «материнской звезды»). В этот момент они хорошо различимы. Надщепление хромосом на хроматиды доходит до центромеры (центральная перетяжка хромосом). Нити веретена деления прикрепляются с одной стороны к центромерам хромосом, с другой стороны - к центриолям. Кариотип организма изучают именно на предмете метафазной клетки. Генетическая характеристика 2n4c.
етафаза. Хромосомы располагаются в экваториальной плоскости клетки (стадия «материнской звезды»). В этот момент они хорошо различимы. Надщепление хромосом на хроматиды доходит до центромеры (центральная перетяжка хромосом). Нити веретена деления прикрепляются с одной стороны к центромерам хромосом, с другой стороны - к центриолям. Кариотип организма изучают именно на предмете метафазной клетки. Генетическая характеристика 2n4c.
Анафаза. Хроматиды начинают расходится к полюсам клетки, двигаясь центромерами вперед, благодаря нитям веретена деления. Укорочение нитей веретена деления осуществляется за счет выпадения участков тубулина. В осуществлении процессов расхождения хромосом важную роль играют компоненты цитоскелета: микротрубочки и микрофиламенты. Генетическая характеристика 4n4c (на полюсах по 2n2c).
Телофаза. Образуется вокруг хромосом ядерная оболочка. Происходит раскручивание хромосом и образование эу- и гегерохроматина, формируется ядрышко.
В конце телофазы происходит деление цитоплазмы - цитотомия или цитокинез, который осуществляется путем образования внутриклеточной пластинки у растительных клеток или путем образования борозды деления у животных клеток. В результате последней фазы митоза образуются две клетки, каждая из которых имеет полный диплоидный набор: 2n2c.
Продолжительность митоза неодинакова в разных типах клеток. В животных клетках среднее время митоза составляет 30-60 минут, а в растительных может достигать 2-3 часов. В жизненном цикле клеток митоз занимает всего 5-10% времени клеточного цикла, все остальное время занимает интерфаза, причем пресинтетический период занимает 30-40%, синтетический 30-50%, постсинтетический - 10-20% времени клеточного цикла. Продолжительность митоза зависит также от размеров клеток, числа ядер, от условий окружающей среды, в частности от температуры.
В организме многоклеточных животных митоз контролируется нервной системой, гормонами надпочечников, гипофиза, щитовидной железы, половых желез и кейлонами. Нарушение нормального взаимодействия различных регуляторных механизмов вызывает изменение митотической активности. Например, в опухолевых клетках митоз выходит из-под контроля нейрогуморальной регуляции. Митоз регулируется, кроме того, взаимодействием организма и среды, то есть имеют значение суточные (циркадные) ритмы. В большинстве органов у животных, активных в ночное время, максимальная частота митозов отмечается утром, а минимальная частота - ночью. У животных, активных днем, и у человека отмечается обратная картина митотической активности.
Таким образом, биологическое значение митоза состоит в обеспечении клеток равноценной наследственной информацией и равномерном распределение хромосом между дочерними клетками.
Митоз происходит в соматических клетках, это все клетки организма, кроме половых.
Известны и некоторые другие способы репродукции клеток - эндомитоз, политения и амитоз. При эндомитозе и политении не происходит ни образования новых ядер, ни деления клетки. При эндомитозе происходит увеличение числа хромосом, т.е. возникают полиплоидные клетки, и нарастает масса цитоплазмы. Политения заключается в образовании крупных хромосом, что также ведет к увеличению массы цитоплазмы и ядра.
3.6.2. Амитоз
А митоз - прямое деление ядра, то есть деление клеточного ядра без образования хромосом и веретена деления. В результате амитоза образуется два или несколько дочерних ядер, причем они могут быть как равноценны, так и неравноценны по содержанию хромосом. Вслед за делением ядра происходит деление цитоплазмы. Если цитоплазма не делится, образуются многоядерные клетки. Амитоз характерен для высокодифференцированных клеток (нейроны, хрящевые, железистые клетки, лейкоциты крови, а также клетки злокачественных опухолей). Вопрос о биологическом значении амитоза еще не решен. Однако не возникает сомнения, что амитоз - вторичное явление по отношению к митозу, поскольку ядерная мембрана и ядрышко сохраняются, а само ядро продолжает активно функционировать.
митоз - прямое деление ядра, то есть деление клеточного ядра без образования хромосом и веретена деления. В результате амитоза образуется два или несколько дочерних ядер, причем они могут быть как равноценны, так и неравноценны по содержанию хромосом. Вслед за делением ядра происходит деление цитоплазмы. Если цитоплазма не делится, образуются многоядерные клетки. Амитоз характерен для высокодифференцированных клеток (нейроны, хрящевые, железистые клетки, лейкоциты крови, а также клетки злокачественных опухолей). Вопрос о биологическом значении амитоза еще не решен. Однако не возникает сомнения, что амитоз - вторичное явление по отношению к митозу, поскольку ядерная мембрана и ядрышко сохраняются, а само ядро продолжает активно функционировать.
РЕКОМЕНДАЦИИ ДЛЯ САМОПОДГОТОВКИ
При подготовке к практическому занятию по материалам учебного модуля вспомните, с участием каких органелл в клетке происходят следующие процессы: синтез белка; синтез нуклеиновых кислот; синтез жиров и углеводов; фотосинтез; гликолиз; биологическое окисление.
Составьте схему биосинтеза белка с молекулы ДНК следующей последовательности:
АТТ .ТТЦ. ГГА. АГА. ацц. атц. ТТЦ. ГГА. АГА.
Составьте схему последовательных реакций фотосинтеза.
Составьте схему последовательных реакций биологического окисления.
тест для проверки знаний
Дополните
Совокупность реакций биосинтеза называется ___________________________________________
Совокупность реакций расщепления называется _________________________________________
Расщепление белков идет до ____________________________________________________________
Расщепление жиров идет до ____________________________________________________________
Расщепление углеводов идет до _________________________________________________________
Обмен веществ идет в __________________________________ этапа.
Первый этап обмена веществ называется________________ и происходит в __________________
Второй этап обмена веществ называется_________________________________________________
Бескислородный этап энергетического обмена называется ________________________________
Кислородный этап энергетического обмена происходит в _________________________________
Суммарное уравнение реакций гликолиза ________________________________________________
Суммарное уравнение реакций расщепления глюкозы ____________________________________
Суммарное уравнение реакций фотосинтеза ______________________________________________
Суммарное уравнение реакций темновой фазы фотосинтеза________________________________
Кислород при фотосинтезе образуется за счет _____________________________________________
Процесс синтеза и-РНК называется _____________________________________________________
Процесс редактирования и-РНК называется _____________________________________________
Процесс синтеза белка на рибосомах называется _________________________________________
Процессы транскрипции в эукариотической клетке осуществляются в _____________________
Стадии транскрипции__________________________________________________________________
Стадии трансляции__________________________________________________________________
Фермент, обеспечивающий процесс транскрипции ________________________________________
Выберите номер правильного ответа
Кодон - это -
триплет ДНК
триплет и-РНК
триплет ДНК, и РНК, РНК вирусов
триплет т-РНК
24. Синтез АТФ в клетке осуществляется в
цитоплазме
митохондриях
ЭПС
митохондриях и цитоплазме
25. Реакции биосинтеза в клетке идут с
поглощением энергии
выделением энергии
26. Энергия, образовавшаяся на подготовительном этапе энергетического обмена
расходуется на синтез АТФ
расходуется на биосинтез различных веществ
не запасается, а рассеивается в виде тепла
Выберите номера всех правильных ответов
27. Автотрофами являются
грибы
сине-зеленые водоросли
зеленые растения
эвглена зеленая
28. Гетеротрофами являются
бактерии
водоросли
сине- зеленые водоросли
плоские черви
грибы
29. Хемосинтезирующими организмами являются
сине-зеленые водоросли
зеленые растения
метанообразующие бактерии
тионовые бактерии
Выберите номер правильного ответа
30. Световая фаза фотосинтеза идет
в строме хлоропластов
на мембранах тилакоидов
внутри тилакоидов гран
Выберите номер правильного ответа
31. Хлорофилл поглощает лучи спектра
красные
зеленые
фиолетовые
32. Темновая фаза фотосинтеза идет
в строме хлоропластов
на мембранах тилакоидов
внутри тилакоидов гран
33. Наибольшее количество энергии выделяется в процессе
брожения
фотосинтеза
дыхания
34. Процессы аэробного окисления протекают в
митохондриях
пластидах
цитоплазме
рибосомах
Задачи для Проверки знаний и умений
Задача 1
Бета-лактоглобулин - белок коровьего молока имеет генетические варианты А,В и с, различающиеся по аминокислотам в позициях В и С. На других участках аминокислоты, входящие в состав белка, одинаковые.
Аминокислотный состав в этих позициях следующий:
ала вариант В
Ала – глу – про – глу - глн - сер – лей - - цис
вал вариант С
позиции 111 – 112 – 113 – 114- 115 - 116 - 117 - 118 - 119
Определите последовательность нуклеотидов в матричной и комплиментарной ей нитях молекулы ДНК в варианте В.
Какие изменения произошли в последовательности нуклеотидов в молекуле ДНК в варианте С?
Определите антикодоны в т-РНК.
Задача 2Установите длину гена, кодирующего синтез белка коровьего молока бета-лактоглобулина, если известно, что он состоит из 350 аминокислот, а расстояние между нуклеотидами ДНК составляет 0, 34 нм.
Найдите молекулярную массу гена, если средняя молекулярная масса нуклеотида равна 340 дальтон.
Задача 3.
В одной из цепочек молекулы ДНК нуклеотиды расположены в такой последовательности: АЦГ ТТА ГЦТ АГТ … Какова последовательность нуклеотидов в другой цепочке белка этой же молекулы? Какой аминокислотный состав имеет белок, кодируемый данной ДНК?
Длина гена, контролирующего синтез белка составляет 335,24 нм. Определите, сколько аминокислот входит в состав белка, если расстояние между двумя нуклеотидами составляет 3,4 А0.. Найдите молекулярную массу гена, если средняя молекулярная масса нуклеотида равна 340 дальтон.
Укажите последовательность аминокислот в белковой молекуле, кодируемой ДНК:
.. АТА ЦТГ АЦА ТТА ГАА
Какой будет последовательность аминокислот, если между 10 и 11 нуклеотидом произойдет вставка гуанина?
Длина гена, контролирующего синтез белка составляет 7799,6 А0. Определите, сколько аминокислот входит в состав белка, если расстояние между двумя нуклеотидами составляет 3,4 А0 Найдите молекулярную массу гена, если средняя молекулярная масса нуклеотида равна 340 дальтон.
Выпишите нуклеотиды м-РНК, кодирующие аминокислотный состав белковой молекулы в следующих вариантах:
а) аспарагин – аланин – тирозин- лизин;
б) фенилаланин –изолейцин- валин-глицин;
Одна цепь участка ДНК имеет следующую последовательность оснований:
ГТАГЦЦТАЦЦЦАТАГГ 3
а) Допустим, что с этой ДНК транскрибируется м-РНК, причем матрицей служит комплиментарная цепь. Какова будет последовательность и-РНК?
б) Сколько пептидов кодирует эта и-- РНК?
Одна цепь участка ДНК имеет следующую последовательность оснований:
ГТАГЦЦТАЦЦЦАТАГГ 3
Допустим, что с этой ДНК транскрибируется м-РНК, какова будет последовательность и-РНК?
При неполном окислении 38 молекул глюкозы образовалось 18 молекул молочной кислоты. Сколько молекул АТФ, кислорода и углекислого газа образовалось при окислении глюкозы?
Дополнительная литература
О.В. Быков В.Л. Цитология и общая гистология (функциональная морфология клеток и тканей человека). – СПб.: СОТИС, 1998.
Джавец Э., Мельник ДЖ.Л., Эйдельберг Э.Руководство по медицинской микробиологии: – М.: Медицина, 1982
Леви А., Сикевиц Ф. Структура и функции клетки. – М.: Мир, 1971.
Международная гистологическая номенклатура / Под. ред. В.В.Семченко, Р.П. Самусева, М.В.Моисеева и З.Л. Колосовой. – Омск: Омская медицинская академия, 1999
Рейва П., Эверт Р., Айкхрн С. Современная ботаника: в 2-х т. Т.1.: - М., Мир, 1990
Ченцов Ю.С. Общая цитология. – М.: Изд-во МГУ, 1995
Ченцов Ю.С. Введение в клеточную биологию. – М.: ИКЦ «Академкнига», 2004
Хэм А., Кормак Д. Гистология: - М., Мир, 1982 –Т.1.
Шлегель Г. Общая микробиология:- М., Мир, 1987
Якушкин Физиология растений: - М. Просвещение, 1993
93
Просмотр содержимого документа
«клетка модуль9»
Раздел 4. основы патологии клетки
При изучении этого раздела Вы узнаете:
сущность понятий некроз и апоптоз;
причины патологических изменений в клетках;
причины и механизм старения клетки;
механизм и причины некротических изменений клетки;
механизм и этапы апоптоза;
биологическое значение апоптоза.
ИНФОРМАЦИОННЫЙ БЛОК
4.1. Гибель клетокНа воздействие различных видов стресса (повышение температуры, угнетение энергетического обмена, заражение вирусами, нехватка кислорода или глюкозы, повреждение окислителями, химическими препаратами, тяжелыми металлами и др.) все клетки, в том числе и клетки млекопитающих и человека, отвечают стереотипной реакций, охватывающей ядерный аппарат и компоненты цитоплазмы. В основе этой реакции лежит резкое изменение характера экспрессии генов. Она проявляется усилением синтеза особой группы защитных стрессовых белков при подавлении продукции остальных.
Стрессорные белки первоначально были обнаружены при изучении реакции клетки на повышение температуры, поэтому их назвали белками теплового шока, или HSP ( сокращение от анг. Heat Shock Proteins). В дальнейшем был установлен их универсальный характер. HSP действуют на уровне ядра и отдельных компонентов цитоплазмы, они выполняют роль молекулярных спутников структурных белков клетки, обеспечивая их сборку и поддержание нативной конформации, взаимодействие между собой и направленный транспорт. Белки теплового шока предотвращают агрегацию белков и их дальнейшее повреждение в условиях нарушения метаболизма клетки.
4.1.1. Старение клеток
После функционирования в течении определенного периода времени клетка гибнет, при чем ее гибели предшествует период старения. При старении клетка утрачивает способность к репликации ДНК и задерживается в пресинтетическом периоде интерфазы, переходя в G0 – фазу, в отличие от нормальной покоящейся клетки на нее не действуют митогены. Механизмы и смысл клеточного старения остается предметом дискуссии. Согласно одной гипотезе, клеточное старение – результат катастрофического накопления ошибок биосинтетических механизмов клетки, согласно другой – оно является способом защиты организма от рака путем ограничения возможностей роста клеток. Возможно, старение клеток служит механизмом стабилизации размеров взрослого организма.
Морфологические признаки старения клетки: уменьшение ее объема, редукция большинства органелл, увеличение числа крупных лизосом, накопление пигментных и жировых включений, нарастание проницаемости клеточных мембран, вакуализация цитоплазмы.
Гибель клеток, наряду с их размножением и дифференцировкой, является одним из ключевых процессов и факторов в обеспечении нормальной жизнедеятельности различных тканей. Смерть отдельных клеток или их групп постоянно встречается у многоклеточных организмов, так же как гибель одноклеточных организмов. Причины гибели, процессы морфологического и биохимического характера развития клеточной смерти могут быть различными. Можно четко разделить на две категории: некроз (от греч. nekrosis – омертвление) и апоптоз (от греч. корней, означающих «отпадение» или «распадение»), который часто называют программируемой клеточной смертью или даже клеточным самоубийством.
4.1.2. Некроз
Некроз возникает под действием резковыраженных повреждающих факторов – перегревания (гипертермии), переохлаждения (гипотермии), нехватка кислорода (гипоксии), нарушения кровоснабжения (ишемии), метаболических ядов, химических препаратов, механической травмы и др. Некроз представляет собой «смерть в результате несчастного случая» и часто охватывают различные по численности группы клеток.
Структурно-функциональные изменения клеток при некрозе на начальных стадиях его развития проявляются набуханием цитоплазмы и отдельных органелл (в особенности, митохондрий). Отмечается дисперсия рибосом, расширение цистерн ЭПС. Эти изменения связаны с нарушением избирательной проницаемости клеточных мембран, и развиваются в ответ на прекращение деятельности мембранных насосов (из-за непосредственного действия ядов или отсутствия необходимой энергии). При повышении проницаемости плазматической мембраны клетка набухает за счет ее обводнения, в цитоплазме происходит увеличение концентрации ионов Na+ и Ca2+, закисление цитоплазмы, набухание вакуолярных компонентов и разрыв их мембран, прекращение синтеза белков в цитозоле, освобождение лизосомных ферментов-гидролаз и лизис клетки. Одновременно с этими изменениями в цитоплазме изменяются и клеточные ядра: вначале они компактизируются (пикноз ядер), но по мере набухания ядра и разрыва его оболочки пограничный слой хроматина распадается на мелкие массы (кариорексис) а затем наступает кариолизис, растворение ядра.
Особенностью некроза является то, что такой гибели подвергаются большие группы клеток (например, при инфаркте миокарда из-за прекращения снабжения кислородом участка сердечной мышцы). Обычным является то, что участок некроза подвергается атаке лейкоцитов и в зоне некроза развивается воспалительная реакция.
4.1. 3. Апоптоз
В процессе развития организмов и их функционировании во взрослом состоянии постоянно происходит гибель части клеток, но без их физического или химического повреждения, происходит как бы их «беспричинная» смерть. Гибель клеток происходит практически на всех стадиях онтогенеза. Многочисленны примеры отмирания клеток без повреждения при эмбриональном развитии. Так отмирают клетки вольфова и мюллерова каналов при развитии мочеполовой системы у позвоночных, погибает часть нейробластов и гонадоцитов, погибают клетки при метаморфозах насекомых и амфибий (резорбция хвоста у головастика и жабр у тритона) и т.д.
Во взрослом организме также постоянно происходит «спонтанная» гибель клеток. Миллионами погибают клетки крови нейтрофилы, клетки эпидермиса кожи, клетки тонкого кишечника - энтероциты. Погибают фолликулярные клетки яичника после овуляции, погибают клетки молочной железы после лактации. Особенно много примеров гибели клеток без непосредственного их повреждения при различных патологических процессах. Например, кастрация (удаление семенников) вызывает гибель клеток предстательной железы, удаление гипофиза приводит к гибели клеток надпочечников. Другой пример - гибель шванновских клеток при дегенерации аксона. Эти клетки в поврежденном периферическом нерве взрослого животного, так же как и клетки-сателлиты и чувствительные нейроны в соответствующих спинномозговых узлах, погибают.
Эти наблюдения наводят на мысль, что клеточная смерть регулируется межклеточными взаимодействиями различным образом. Множество клеток многоклеточного организма нуждается в сигналах на то, чтобы оставаться живыми. В отсутствии таких сигналов или трофических факторов в клетках развивается программа «самоубийства» или программируемой смерти. Например, клетки культуры нейронов погибают при отсутствии фактора роста нейронов (NGF), клетки простаты гибнут в отсутствии андрогенов семенника, клетки молочной железы погибают при падении уровня гормона прогестерона и т.д. С другой стороны, клетки могут получать сигналы, которые в клетках-мишенях запускают процессы, приводящие к гибели по типу апоптоза. Так, гидрокортизон вызывает гибель лимфоцитов, а глютамат - нервных клеток в культуре ткани, фактор некроза опухоли (TNF) вызывает гибель самых различных клеток. Тироксин (гормон щитовидной железы) вызывает апоптоз клеток хвоста головастиков. Кроме этого существуют ситуации, когда апоптическая гибель клетки вызывается внешними факторами, например, радиацией.
Понятие «апоптоз» было получено при изучении гибели части клеток печени при неполной перевязке портальной вены. Было обнаружено, что при этом наблюдается своеобразная картина клеточной смерти, которая затрагивает лишь отдельные клетки в паренхиме печени. Процесс начинается с того, что клетки теряют контакты с соседними клетками, они как бы сморщиваются (первоначальное название этой формы гибели shrinkage necrosis – некроз сжатием клетки). На электронно-микроскопическом уровне выявляется утрата клетками специализированных структур на их поверхности микроворсинок, межклеточных соединений. В ядрах по их периферии происходит специфическая конденсация хроматина, затем ядро фрагментируется на отдельные части, вслед за этим сама клетка фрагментируется на отдельные тельца, отграниченные плазматической мембраной – апоптозные тела. Апоптоз - процесс, приводящий не к лизису, не к растворению клетки, а к ее фрагментации, распаду. При апоптозе цитоплазматические структуры апоптических телец сохраняют свою целостность.
И
зменение в ядре при апоптозе обусловлены активизацией эндогенной Ca2+ Mg2+-зависимой эндонуклеазы, что приводит к упорядоченному расщеплению геномной ДНК в межнуклеосомных участках на отдельные нуклеосомные сегменты. Хроматин укладывается в ядре в виде крупных полулуний, после чего ядро распадается на фрагменты, окруженные мембраной. Таким образом, в отличие от некроза, изменения ядра при апоптозе включают только пикноз и своеобразный кариорексис (без разрушения ядерной оболочки), кариолизис отсутствует
При прогрессировании апоптоза нарастающая конденсация цитоплазмы сочетается с изменением формы клетки – она образует многочисленные крупные вздутия и выпячивания, а также «кратеры» - клетка как бы вскипает.
Выпячивания, содержащие целостные органеллы, а также фрагменты ядра, отшнуровываются, формируя апоптозные тела. Образование апоптозных тел связано с изменениями цитоскелета, в частности активизацией актиновых микрофиламентов. Судьба апоптических телец тоже необычна: они фагоцитируются макрофагами или даже нормальными соседними клетками. При этом не развивается воспалительная реакция. Апоптозный процесс развивается очень быстро и обычно длится от нескольких минут до нескольких часов (в среднем не более 1-3 ч.)
Сигнал, запускающий апоптоз, инициирует внутриклеточную каталитическую реакцию, которая включает ряд этапов:
передача сигнала в клеточное ядро, которая обеспечивается различными механизмами, опосредованными ионами Ca2+, фосфолипазой, тирозинкиназой, протеинкиназами А, С и другими молекулами;
активизация «летальных» или «киллерных» генов, ответственных за развертывание программы апоптоза;
включение процессов транскрипции и трансляции синтеза аппоптоз-специфических белков;
активизация ферментативных систем, вызывающих необратимые изменения в клетке.
Важно отметить, что во всех случаях апоптоза, во время ли эмбрионального развития, во взрослом ли организме, в норме или при патологических процессах, морфология процесса гибели клеток очень сходна. Это может говорить об общности процессов апоптоза в разных организмах и в разных органах.
Исследования на разных объектах показали, что апоптоз есть результат реализации генетически запрограммированной клеточной гибели.
Апоптоз – один из фундаментальных и универсальных механизмов тканевого гомеостаза.
Особенно значима роль апоптоза в следующих процессах.
Эмбриональное развитие сопровождается избыточным образованием большого числа клеток, которые своевременно уничтожаются апоптозом. Наиболее активно этот процесс происходит в нервной системе, где механизмом апоптоза гибнет до 40 –85% клеток различных участков ЦНС. Процессы образования просветов в трубчатых органов (сосудов, кишечника, желудка, трахеи, бронхов и др.), инволюции провизорных органов (амнион, желточный мешок, аллантоис), разрыв плодных оболочек осуществляются путем апоптоза. Одним из механизмов действия тератогенных факторов (от греч. Teras – урод и genos – происшедший), веществ, обуславливающих развитие уродств, - изменение расположения зон, в которых в норме происходит гибель клеток путем апоптоза.
Апоптоз стареющих клеток в зрелых тканях. Старение клеток в физиологических кусловиях завершается апоптозом. Развитие апоптоза индуцируется накоплением генетических ошибок и (или) снижением чувствительности клетки к стимулирующим ростовым сигналам.
Апоптоз при инволюции зрелых тканей особенно выражен в гормонально-зависимых органах после прекращения их гормональной стимуляции. Он характерен для органов половой системы после удаления гонад, для постлактационной инволюции молочной железы, послеродовой инволюции матки.
А поптоз в клетках иммунной системы обеспечивает развитие и течение всех важнейших иммунных реакций.
поптоз в клетках иммунной системы обеспечивает развитие и течение всех важнейших иммунных реакций.
Апоптоз в реакции тканей на повреждающее действие различных факторов, таких как радиация, действие некоторых токсинов, ингибиторов клеточного метаболизма. Многие токсины (рицин, дифтерийный токсин и др.), а также антиметаболиты могут вызывать гибель клеток путем апоптоза
Апоптоз имеет значение и в развитии дегенеративных и инфекционных заболеваний. Патологическая активизация апоптоза может играть роль в развитии таких заболеваний, как болезнь Альцгеймера, Паркинсона, Гентингтона Лу-Ринга. Апоптоз может запускаться в клетках человека и при инфицировании бактериями или вирусами, в частности при ВИЧ-инфицировании.
Угнетение апоптоза может служить одним из механизмов канцерогенеза – развития опухолей.
Элиминация, удаление отдельных клеток путем апоптоза наблюдается и у растений.
Биологическая роль апоптоза или программированной смерти клеток очень велика: это удаление отработавших свое или ненужных на данном этапе развития клеток, а также удаление измененных или патологических клеток, особенно мутантных или зараженных вирусами.
Таким образом, апоптоз – это генетически запрограммированная гибель клеток, которая приводит к «аккуратной» разборке и удалению клеток.
РЕКОМЕНДАЦИИ ДЛЯ САМОПОДГОТОВКИ
Вопросы для обсуждения и размышления
Многие так называемые биологически активные добавки содержат вещества, активизирующие процессы жизнедеятельности клеток организма. Насколько оправдано использование этих БАДов без консультации врача особенно в климактерический период.
В нетрадиционной медицине используется перекись водорода для внутреннего применения. Перекись водорода является сильнейшим окислителем. Предположите последствия использования такого «лечения».
Известно, что лучевая энергия является сильнейшим мутагеном. Активному солнечному излучению летом организм человека «готовиться» благодаря сложнейшим механизмам активизации синтеза меланина в структурах кожи. Почему во многих странах ограничено использование соляриев?
Известный постулат Р.Вирхова гласит: «…любая патология организма начинается с патологии клетки». Перечислите клеточные механизмы защиты организма от развития патологии.
Насколько правомерна реклама «… препарат подходит к каждой клетке и восстанавливает здоровье на клеточном уровне»? Какими свойствами должно обладать данное вещество? Какую химическую природу иметь? Как можно ввести этот препарат, если мы знаем, что полимеры расщепляются уже на подготовительном этапе обмена веществ?
В чем значение белков теплового шока? Как они защищают клеточные структуры?
Темы учебно-исследовательских работ
Биологическое значение апоптоза.
Почему некроз вызывает воспалительный процесс?
Проблемы и гипотезы старения.
Старость. Это благо или зло?
Как можно продлить жизнь клеток и организма в целом?
Дополнительная литература
О.В. Быков В.Л. Цитология и общая гистология (функциональная морфология клеток и тканей человека). – СПб.: СОТИС, 1998.
. Леви А., Сикевиц Ф. Структура и функции клетки. – М.: Мир, 1971.
Меркулов Г.А. Курс патологогистологической техники. – Изд-во Медицина, Ленинградское отделение, 1969.
Ченцов Ю.С. Общая цитология. – М.: Изд-во МГУ, 1995
Ченцов Ю.С. Введение в клеточную биологию. – М.: ИКЦ «Академкнига», 2004
Щелкунов С.И. Клеточная теория и учение о тканях. – Медгиз, Ленинградское отделение, 1958.
СЛОВАРЬ ЦИТОЛОГИЧЕСКИХ ТЕРМИНОВ
АДЕНОЗИНТРИФОСФАТ (АТФ) — аккумулятор и источник энергии в организмах. АТФ выполняет функцию поставщика энергии, переноса одной из своих богатых энергией фосфатных групп на другую молекулу, в результате чего АТФ превращается в аденозиндифосфат (АДФ). В митохондриях АДФ «перезаряжается», присоединяя к себе фосфатную группу, и вновь превращается в АТФ. Высвобожденная из АТФ энергия трансформируется в другие виды энергии, необходимые для различных физиологических процессов (биосинтез высокомолекулярных соединений, мышечное сокращение, нервная деятельность и др.).
АДЕНОЗИНТРИФОСФАТАЗА - фермент, катализирующий гидролиз аденозинтрифосфорной кислоты (АТФ). При гидролизе от нее отщепляется один остаток фосфорной кислоты с образованием аденозиндифосфорной (АДФ) и фосфорной кислот. За счет разрыва АТФ освобождается энергия, используемая организмом для различных физиологических функций.
АКТИН - белок микрофиламентов мышечных и других клеток. А. соединяется с миозином, образуя актомиозин, обусловливающий сократительную способность мышц.
АМИТОЗ - изредка встречающийся тип клеточного деления, при котором сохраняется внутренняя структура ядра (хромосомы не выявляются, и веретено деления не образуется).
АНАФАЗА - фаза расхождения парных хромосом в процессе клеточного деления - митоза и мейоза.
АНГСТРЕМ (А) - единица длины, равная 1010 м.
АНИМАЛЬНЫЙ - животный, относящийся к животному.
АНТИГЕН - чужое для организма вещество (обычно белкового характера), вызывающее в теле животных и человека образование антител. См. Антитела.
АНТИТЕЛА - противотела, белковые вещества (в большинстве гамма-глобулины), вырабатываемые лимфоцитами и плазматическими клетками. А. вступают в нейтрализирующую или разрушающую реакцию только с антигенами, вызывающими их продукцию. В соответствии с образом действия Антитела распадаются на агглютинины, бактериолизины, гемолизины, преципитины и др. Антитела накапливаются в сыворотке крови и тканях. См. Антигены.
АПИКАЛЬНЫЙ - верхушечный, обращенный кверху.
АПОПТОЗ – генетически запрограммированная смерть клетки.
АРГЕНТАФИННАЯ КЛЕТКА - клетки, окрашивающиеся солями серебра и хрома и содержащие гранулы, способные восстанавливать из солей металлическое серебро.
АРГИРОФИЛЬНАЯ КЛЕТКА - клетка, которая импрегнируется серебром только при применении соответствующего восстановителя.
АТРОФИЯ - уменьшение объема ткани или органа в результате общего или местного нарушения питания. Атрофия бывает физиологическая и патологическая. Развивается атрофия медленно и является хроническим процессом.
АУТОЛИЗ - посмертное растворение клеток и тканей под действием собственных ферментов.
АУТОРАДИОГРАФИЯ - метод демонстрации определенных химических веществ путем мечения их радиоактивными изотопами (Н3, С14, Р32 и др.) и последующего выявления их излучения на фотографической пластинке.
АУТОФАГОСОМА - лизосома, содержащая изношенные и переваренные в разной степени клеточные органеллы.
АЦИДОФИЛИЯ - свойство клеток окрашиваться кислыми красителями (в противоположность базофилии - окрашивании основными красителями).
БАЗАЛЬНАЯ МЕМБРАНА - внеклеточный конденсированный слой гликопротеина, мукополисахарида и белков, встречающийся под базальной поверхностью любых эпителиев. Базальная мембрана состоит из плотного филаментозного листка, базального слоя и из расположенного глубже ретикулярного слоя.
БАЗАЛЬНОЕ ТЕЛЬЦЕ - модифицированный центриоль, расположенный между основанием ресницы и ее корешком.
БАЗОФИЛИЯ - свойство структурных компонентов клетки окрашиваться основными красителями. Интенсивная базофиия хроматина зависит от наличия в ней дезоксирибонуклеиновой кислоты, а в цитоплазме базофильными являются структуры, содержащие рибонуклеиновую кислоту, мукополисахариды, белки и другие соединения. Базофилия характерна для активно растущих, интенсивно синтезирующих белок клеток.
БИВАЛЕНТ - обе гомологичные хромосомы диплоидного организма, конъюгирующие в мейозе. В норме число бивалентов равно числу хромосом гаплоидного набора.
БОКАЛОВИДНЫЙ ЭКЗОКРИНОЦИТ - одноклеточная железа, выделяющая слизь; встречается в эпителии слизистых оболочек дыхательных путей и кишечника.
ВАКУОЛЯ - внутриклеточная шаровидная полость, окруженная мембраной и содержащая жидкость, или плотное вещество, или то и другое вместе.
ВЕЗИКУЛА - пузырьковидная структура, содержащая жидкость.
ВИТАЛЬНОЕ ОКРАШИВАНИЕ - окрашивание живых организмов или их частей (тканей, клеток) в целях их изучения; прижизненное окрашивание.
В-ЛИМФОЦИТЫ - тимус-независимые лимфоциты; они поступают из костного мозга в ткани, не проходя тимуса и не испытывая его влияния. В-лимфоциты аналогичны лимфоцитам, продуцируемым у птиц в клоакальной сумке (бурса-эквивалентные лимфоциты). В-лимфоциты, созревая, превращаются в плазмоциты, синтезирующие антитела.
ГАМЕТА - гаметы - половые клетки,- сперматоциты и овоциты на всем протяжении их развития.
ГАПЛОИДНОЕ ЧИСЛО ХРОМОСОМ - половинное число хромосом, образующееся в зрелых половых клетках (спермиях и яйцах) в результате редукционного деления.
ГЕМОГЛОБИН - сложный белок из группы хромопротеидов, содержащийся в эритроцитах. Обладает свойством легко окисляться и быстро восстанавливаться, отдавая кислород клеткам.
ГЕН - элементарная единица наследственности, представленная специфическими участками (локусами), дифференцированными по длине хромосом.
ГЕНЕРАЦИЯ - период жизни организма или клетки от начала их развития до половозрелого состояния или до нового размножения (митоза).
ГЕТЕРОГАМЕТНЫЙ - организм, продуцирующий половые клетки с неравным числом половых хромосом (у млекопитающих мужской пол).
ГИПЕР - приставка, означающая усиление основного понятия, излишек, над, сверх, слишком; соответствует латинским приставкам зирег и рег.
ГИПЕРТРОФИЯ - увеличение объема ткани или органа с сохранением обычных пропорций и форм. Термин гипертрофия. применяется лишь к местным изменениям, к которым не относятся возрастные увеличения и увеличения болезнетворного происхождения.
ГИПО - приставка, обозначающая уменьшение, ослабление основного понятия, уменьшение ниже нормы, несовершенство.
ГЛИКОКАЛИКС - гликопротеидный материал тонкофибриллярного характера, покрывающий поверхность всех клеток, независимо от того, соприкасаются они с поверхностями других клеток или являются свободными. Гликокаликс очень тесно связан с плазмалеммой (поверхностный слой плазмалеммы).
ГОЛЬДЖИЕВЫЙ КОМПЛЕКС - клеточная органелла, состоящая из нескольких пакетов двойных мембран, вакуолей и мешочков. Комплекс Гольджи является местом накопления и концентрирования секреторного белка и его упаковки в мембранные везикулы. Комплекс Гольджи участвует также в биосинтезе липо- и гликопротеинов, фосфолипидов и мембранных протеинов.
ГОРМОНЫ - органические биологически высокоактивные вещества, образуемые эндокринными железами (а также отдельными клетками с эндокринной функцией) и выделяемые ими непосредственно в кровь и лимфу. По своей химической структуре гормоны подразделяются на стероидные (половые гормоны и гормоны надпочечников), белковые и пептидные (гормоны, вырабатываемые гипофизом, щитовидной, паращитовидной железами, панкреасом и мозговым слоем надпочечников). По их физиологическому действию гормоны. распадаются: на кинетические гормоны, оказывающие сравнительно кратковременное действие на некоторые гладкомышечные структуры и железы (окситоцин, вазопрессин, адреналин и норадреналин); метаболические гормоны, участвующие в регуляции обменных процессов (тироксин, кальцитонин, паратгормон, инсулин, глюкагон), и на морфогенетические гормоны, участвующие в контроле роста и дифференциации клеток, тканей и органов (соматотропный гормон, фолликулостимулирующий гормон, эстрогены, тестостерон) .
ДЕДИФФЕРЕНЦИАЦИЯ - потеря характерных признаков у специализированных клеток и их возвращение к незрелому состоянию.
ДЕСМОСОМА - двураздельная дискообразная структура диаметром около 200—400 нм, образованная утолщенной плазматической мембраной двух смежных клеток и более толстым слоем цитоплазматических фибрилл.
ДИАСТЕР - двойная звезда, две группы хромосом, расположенных на полюсах митотического веретена в анафазе митоза.
ДИПЛОИДНЫЙ НАБОР ХРОМОСОМ - наличие в клетках полного набора пар гомологичных хромосом, характерный для зиготы и всех соматических клеток, за исключением анэуплоидных, полиплоидных и гаплоидных клеток.
ДИФФЕРЕНЦИРОВКА КЛЕТОК - развитие клеток, проявляющееся в их превращении из ранее однородных (индифферентных) структур в неоднородные (специализированные) клетки.
ДИФФЕРОН - совокупность по одной линии дифференцированных тканевых клеток, берущих начало от одного вида стволовых клеток.
ЗИГОТА - клетка, возникающая в результате слияния двух гамет (спермия с яйцом) с одинарным (гаплоидным) числом хромосом; оплодотворенное яйцо с удвоенным (диплоидным) числом хромосом.
ЗИМОГЕН - инактивный предшественник энзимы, который превращается в действующую энзиму под влиянием кислоты, другой энзимы или иным способом (проэнзима).
КУЛЬТУРА ТКАНЕЙ - метод выращивания в искусственно созданных условиях фрагментов органов и тканей или популяции отдельных изолированных клеток. В зависимости от характера роста и биологических особенностей эксплантатов, а также методов их приготовления различают следующие типы Культура тканей: в сгустке плазмы, однослойные, суспензионные, органные культуры, культуры переживающих тканей,
ЛИЗОСОМЫ - вид субмикроскопических (0,4 мкм) телец (пищеварительных органелл) в цитоплазме, ограниченных единственной мембраной и содержащих около дюжины гидролитических энзимов. Энзимы лизосом обеспечивают переваривание захваченных клеткой частиц или отмирающих структур цитоплазмы. Лизосомы идентифицируются посредством гистохимического метода на кислую фосфатазу.
ЛИПОПРОТЕИДЫ - комплексы различных белков (глобулинов, альбуминов и др.) с липидами, содержатся во всех тканях организма, больше всего их в нервной ткани.
ЛИПОФУСЦИН - бурый пигмент, пигмент старения, зернистый липопротеид желто-коричневого цвета. Обнаруживается главным образом в старости при истощающих болезнях.
МАКРОСКОПИЧЕСКИЙ - видимый невооруженным глазом (без помощи микроскопа).
МЕЙОЗ - тип двойного ядерного деления, свойственного половым клеткам (сперматоцитам и овоцитам), при котором количество хромосом, характерное для соматических клеток (диплоидное количество - п), уменьшается вдвое (гаплоидное количество - 1п). Профаза первого мейотического деления является крайне продолжительной (2-3 недели) и распадается на пролептотенную, лептотенную, инапсальную, зиготенную, пахитенную и диплотенную фазы и на диакинез. У самцов диакинезу безостановочно следуют дальнейшие стадии первого мейотического деления, а у самок диакинез переходит в длинный период покоя, продолжающийся от нескольких месяцев до нескольких лет.
МЕМБРАНЫ БИОЛОГИЧЕСКИЕ - широкие, метаболически активные пленки. К ним принадлежат клеточная мембрана, или плазмалемма, и внутриклеточные мембраны: ядерная оболочка, или нуклеолемма, эндоплазматическая сеть, гольджиевые мембраны, мембраны митохондрий и мембраны, окружающие пиноцитотические и фагоцитотические вакуоли. Биологические мембраны выполняют крайне различные функции: секрецию белков, синтез жиров, детоксикацию некоторых лекарств, контроль фагоцитоза, респираторную функцию митохондрий и активный транспорт веществ.
МЕТАФАЗА - стадия митоза и мейоза, во время которой хромосомы располагаются в виде экваториальной пластинки, или материнской звезды.
МЕТАЦЕНТРИЧЕСКИЕ ХРОМОСОМЫ - хромосомы, имеющие центромеру в своей середине; центромера разделяет хромосомы на два разных по длине плеча.
МИКРОВОРСИНКИ - пальцевидные выросты цитоплазмы, каждый из которых покрыт клеточной мембраной (плазмолеммой) и выступает с поверхности клетки в просвет органа. Некоторое количество микроворсинок встречается почти на всех клетках организма, хотя они редко бывают такими крупными и распределены так равномерно, как в столбчатых эпителиоцитах кишечных ворсинок. Микроворсинки очень сильно увеличивают поверхность, через которую происходит всасывание питательных веществ в тонком кишечнике.
МИКРОМЕТР (мкм) - тысячная часть миллиметра.
МИКРОТРУБОЧКИ - полые неразветвленные цилиндры диаметром в 21-24 нм и длиной в несколько микрометров. Часто они появляются в группе от 30 или 40 единиц. микротрубочки являются составными частями центриолей и ресничек и образуют ахроматический митотический аппарат. Они легко диспергируются под действием колхицина.
МИКРОФИЛАМЕНТЫ - филаменты диаметром в 5-6 нм, состоят из белка, называемого актином; встречаются, кроме мышечных элементов, в самых разнообразных клетках, участвуя в механизме клеточного сокращения.
МИТОХОНДРИИ - цитоплазматические органеллы, содержащие цепи ферментов, осуществляющих клеточное дыхание. Митохондрии ограничены двумя мембранами - наружной и внутренней. Внутренняя мембрана образует складчатые или трубчатые выступы внутрь митохондрии, называемые кристами. Митохондрии наполнены жидкостью, называемой матриксом. Иногда в матриксе лежат электроплотные гранулы. Диаметр митохондрий составляет в большинстве случаев от 0,4 до 1 мкм, а их число измеряется в некоторых (печеночных) клетках сотнями. Ферменты митохондрий катализируют реакции, необходимые для снабжения клетки важным богатым энергией соединением — аденозинтрифосфатом (АТФ). АТФ выполняет свою фукнцию поставщика энергии, перенося одну из своих богатых энергией концевых фосфатных групп на другую молекулу, в результате чего АТФ превращается в аденозиндифосфат (АДФ). В митохондриях АДФ «перезаряжается», присоединяя к себе фосфатную группу, и вновь превращается в АТФ.
МУЛЬТИВЕЗИКУЛЯРНЫЕ ТЕЛЬЦА - лизосомы, содержащие маленькие пузырьки пиноцитозного происхождения.
НАБОР ХРОМОСОМ - число хромосом в клетке. В половых клетках имеется по одному гаплоидному набору хромосом, в соматических их два. Для нормального развития клетки необходим полный (двойной) набор хромосом.
НЕКСУС - пятно плотного контакта, пронизанное множеством открытых каналов, непосредственно соединяющих цитоплазму двух соседних клеток. Нексусы дают возможность ионам и низкомолекулярным веществам переходить из одной клетки в цитоплазму другой, минуя межклеточное пространство. Нексусы. встречаются между эпителиальными, костными, мышечными и нервными клетками.
НУКЛЕИНОВЫЕ КИСЛОТЫ - высокомолекулярные соединения, содержащиеся во всех биологических объектах. Различают рибонуклеиновые кислоты (РНК) и дезоксирибонуклеиновые кислоты (ДНК). В состав РНК входят: пуриновые основания - аденин и гуанин, пиримидиновые основания - цитозин и урацил, углевод рибоза и остаток фосфорной кислоты. В ДНК урацил заменен тимином, а рибоза - дезоксирибозой. В клетке нуклеиновые кислоты. находятся как в свободном состоянии, так и в виде комплексов с белком - нуклеопротеидов.
ОКСИФИЛЬНЫЙ - окрашивающийся кислыми красителями.
ПИНОЦИТОЗ - процесс поглощения, клеткой жидкости и переноса ее вглубь в виде пузырьков субмикроскопического размера.
ПОЛОВОЙ ХРОМАТИН - гранула уплотненного хроматина (вещества Х-хромосомы), обычно прилегающая к ядрышку или к ядерной мембране. У высших млекопитающих встречается у особей женского пола; у самцов половой хроматин часто отсутствует.
ПРОЛИФЕРАЦИЯ - разрастание тканей организма путем размножения его клеток.
ПРОТИВОТЕЛА - то же, что антитела.
ПРОТОПЛАЗМА - содержимое живой клетки; состоит из нуклеоплазмы (ядра) и цитоплазмы (остальная часть клетки). К основным структурам ядра относятся ядрышко и хромосомы. Некоторые структуры цитоплазмы активно участвуют в жизнедеятельности клетки и имеются во всех клетках. Такие универсальные структуры называются органеллами. К ним относят: митохондрии, гольджиевый комплекс, центриоли, клеточные мембраны, эндоплазматическую сеть, рибосомы и лизосомы. Кроме органелл в протоплазме нередко обнаруживают включения - структуры, встречающиеся лишь на некоторых этапах развития и функционирования клеток (капли жира, гранулы белков и полисахаридов, секреторные гранулы и пигменты).
ПРОФАЗА МИТОЗА - первая стадия митоза, на протяжении которой в ядре делящейся клетки появляются хромосомы и начинается формирование митотического веретена.
ПУЛ ПРОЛИФЕРАЦИОННЫЙ - количество размножающихся клеток по отношению ко всей массе клеток данной популяции.
РЕГЕНЕРАЦИЯ - восстановление утраченных частей организма путем размножения или гиперплазии сохранившихся тканевых элементов. Различают регенерацию физиологическую, репаративную и патологическую. Физиологическая регенерация - восстановление тканевых лементов взамен естественно утраченных клеток. Репаративная регенерация происходит при повреждении частей тела под воздействием вредных факторов. Патологическая регенерация характеризуется отклонениями от нормального восстановительного процесса.
РЕСНИЧКИ - органеллы движения клеток, длиной в 5-10 мкм и толщиной 0,2 мкм. Состоят из вне- и внутриклеточной частей, в центре которых располагается осевой филамент, включенный в плазмалемму. В состав осевого филамента входят: пара центральных микротубул, девять периферических дипломикротубул и базальное тельце. В базальном тельце различают базальный корень и прикрепляющиеся к нему исчерченные структуры - базальные ножки. В базальном корне дипломикротрубочки заменены трипломикробутулами, причем центральная микротрубочка здесь отсутствует.
РИБОНУКЛЕИНОВЫЕ КИСЛОТЫ (РНК) - биополимеры, мономерной единицей которых являются рибонуклеотиды. В состав каждого рибонуклеотида входят пуриновое или пиримидиновое основание, углевод и фосфорная кислота. РНК являются главным химическим компонентом рибосом.
РИБОСОМЫ - сферические гранулы диаметром 15-30 нм, располагающиеся свободно в цитоплазме или прикрепленные к наружной поверхности мембран эндоплазматической сети. Рибосомы являются центрами синтеза белков в цитоплазме, причем на свободных рибосомах осуществляется синтез белков, необходимых для самой клетки, а на фиксированных - синтез специфических белков «на экспорт».
СЕКРЕТ - продукт секреторной (синтетической) деятельности экзокринных желез.
ТЕЛОФАЗА - конечная фаза митотического деления, сопровождаемая делением цитоплазмы.
ТЕЛОЦЕНТРИЧЕСКАЯ ХРОМОСОМА - хромосома с апикально расположенным центромером.
ТЕЛЬЦЕ БАЗАЛЬНОЕ - модифицированный центриоль, расположенный между основанием ресницы и ее корешками.
ТЕОРИЯ КЛЕТОЧНАЯ - обобщение, утверждающее, что оба главных звена живой природы - растения и животные - имеют общий структурный элемент - клетку, гомологичный (одинаково развивающийся) и аналогичный (выполняющий сходные функции) в обоих звеньях органической природы,
ФАГОСОМА - фагоцитозный пузырек, отделенный от плазмолеммы и содержащий корпускулярный внеклеточного происхождения материал - продукт фагоцитоза. Соединяясь с лизосомой, энзимы последнего переваривают материал, вносимый в клетку фагосомами. Соединенный продукт фагосомы с лизосомой называют фаголизосомой.
ФАГОЦИТОЗ - способность особых клеток - фагоцитов (нейтрофильных гранулоцитов, макрофагоцитов) захватывать и переваривать живые и неживые твердые частицы. Фагоцитоз играет большую роль в защитных реакциях организма.
ХРОМАТИДА - одна из двух продольных единиц хромосомы (полухромосома). Xроматиды становятся видимыми между ранней профазой и метафазой митоза и между диплотеной и метафазой второго мейотического деления. По завершении этих стадий хроматиды. называются дочерними хромосомами.
ХРОМАТИН - вещество хромосом; состоит из дезоксирибонуклеиновой кислоты и гистона. Наряду с этим в хроматин содержатся и неосновные белки и рибонуклеиновая кислота. Различают эухроматин и гетерохроматин. В покоящемся ядре эухроматин теряет свое сродство к красителям, гетерохроматин сохраняется сильно окрашенным основными красителями.
ХРОМОСОМЫ - самопроизводящие структуры и носители генов, подвергающиеся во время деления ядра характерным структурным изменениям. Хромосомы становятся видимыми во время митоза и мейоза. Длина хромосом определяется степенью их спирализации, а их морфология выражается в последовательности эу- и гетерохроматиновых сегментов в локализации центромеры и в положении вторичной перетяжки. По положению центромеры различают акроцентрические, субметацентрические и метацентрические хромосомы. Последние являются равноплечими, а в акроцентрических хромосомах центромера локализуется терминально. Каждая хромосома имеет характерную для нее форму. Совокупность всех хромосом в клетке образует хромосомный набор, постоянный для каждого вида организмов. Различают одинарный, или гаплоидный, набор хромосом., который содержит зрелые половые клетки, и двойной, или диплоидный, набор хромосом, содержащий по паре гомологичных (равных по форме, величине и генетическим свойствам) хромосом.
ЦИКЛ КЛЕТОЧНЫЙ - цикл жизни клетки, охватывающий полный проход через интерфазу и митоз. В зависимости от активности синтеза дезоксирибонуклеиновой кислоты (ДНК) интерфаза подразделяется на предупликационную стадию (G1), во время которой синтеза ДНК не происходит, на синтетическую, или дупликационную, стадию (S),— период синтеза ДНК, требующий около 8 ч, и концевую, постдупликационную, стадию (G2). Предупликационная стадия интерфазы имеет разную продолжительность, а постдупликационная стадия и стадии митоза продолжаются только минуты или немногие часы. Митоз подразделяется на про-, мета-, ана- и телофазы.
ЦИСТЕРНА - замкнутое пространство неопределенной формы в клетке или органе, заполненное
жидкостью.
ЦИТОГЕНЕТИКА - раздел цитологии, в задачу которого входит изучение закономерностей наследственности на клеточном уровне.
ЦИТОЛОГИЯ - наука о структуре, развитии и функции клеток. Современная Ц. базируется на комплексном (микроскопическом, субмикроскопическом, цитохимическом и т.д.) изучении клеток, в разные периоды их жизни и жизни организмов.
ЦИТОПЛАЗМА - часть клетки, окружающая ядро. Она содержит органеллы и включения и является местом большинства химических реакций клетки (белковой, жировой и углеводный обмен, клеточное дыхание и различные специфические функции).
ЦИТОСКЕЛЕТ - комплекс внутрицитоплазматических филаментов (тонофиламенты, тонофибриллы, терминальная сеть) и микротрубочек, поддерживающих форму клеток.
ЦИТОСПЕКТРОФОТОМЕТРИЯ - метод изучения химического состава клеток, основанный на избирательном поглощении теми или иными веществами лучей с определенной длиной волны. По интенсивности поглощения света, которая зависит от концентрации вещества, производят количественное определение его в клетке.
ЦИТОХИМИЯ - способ изучения локализации различных химических веществ в клетке и их изменений в процессе клеточного развития, роста и функционирования.
ЩЕЛЕВОЙ КОНТАКТ - см. Нексус.
ЩЕТОЧНАЯ КАЕМКА - слой микроворсинок на свободной поверхности кишечного эпителия. Увеличивает всасывающую поверхность клеток.
ЭЛЕКТРОННЫЙ МИКРОСКОП - прибор, в котором для получения увеличенного изображения используется пучок движущихся в вакууме электронов, фокусируемый электрическими или магнитными полями (электронными линзами). Позволяет визуально изучать частицы во много раз меньшие, чем наблюдаемые в световом микроскопе.
ЭНДОЦИТОЗ - захват клеткой крупных частиц, неспособных к диффузии через плазмолемму; Э. охватывает как фагоцитоз, так и пиноцитоз.
ЭРГАСТОПЛАЗМА - см. Эндоплазматическая сеть.
ЯДРО - внутриклеточная структура генетической детерминации и регуляции белкового обмена. Интерфазное Я. состоит из хроматина, ядрышка, нуклеоплазмы и ядерной оболочки - нуклеолеммы, отделяющей Я. от цитоплазмы. Пористая нуклеолемма состоит из внешней и внутренней мембран, разделенных цистерной нуклеолеммы. Хроматин интерфазных ядер представляет собой хромосомы, которые, однако, теряют в это время свою компактную форму, разрыхляясь. Ядрышко - округлой формы тельце - является производным хромосомы и местом образования рибосомных рибонуклеиновых кислот, на которых происходит синтез полипептидных цепей как в ядре, так и в цитоплазме. В ядрышке различают филаментозную и гранулезную части. Филаменты имеют толщину 6-8 нм, а диаметр гранул - около 15-20 нм. Большинство ядер млекопитающих характеризуется половым диморфизмом, так как у самцов одна из парных половых хромосом (в качестве полового хроматина) остается в интерфазе гетерохроматичной. Половой хроматин представлен наиболее четко в нейтрофильных гранулоцитах, эпителиоцитах ротовой полости и нейронах центральной нервной системы. См. Хроматин.
ЯДРЫШКО - см. Ядро.
Содержание
|
| Введение | 3 |
|
| Претест | 4 |
| 1 | Предмет цитологии. Методы изучения клетки | 6 |
| 1.1. | Основные этапы становления цитологии как науки | 6 |
| 1.2. | Основные положения клеточной теории | 8 |
| 1.3. | Методы клеточной биологии | 14 |
| 2. | Общие принципы структурной организации клетки | 25 |
| 2.1 | Особенности строения и физиологии прокариотической клетки | 27 |
| 2.2. | Общий план строения эукариотической клетки | 30 |
| 2.2.1. | Морфофункциональные особенности клеточных мембран | 31 |
| 2.2.2 | Поверхностный аппарат клетки | 33 |
| 2.2.3. | Цитоплазма | 37 |
| 2.2.3.1. | Гиалоплазма | 37 |
| 2.2.3.2. | Эндоплазматическая сеть | 39 |
| 2.2.3.3. | Рибосомы | 40 |
| 2.2.3.4. | Комплекс Гольджи | 41 |
| 2.2.3.5. | Лизосомы | 43 |
| 2.2.3.6. | Агранулярная ЭПС | 45 |
| 2.2.3.7. | Вакуоли растительных клеток | 46 |
| 2.2.3.8. | Сферосомы | 46 |
| 2.2.3.9. | Пероксисомы | 47 |
| 2.2.3.10. | Митохондрии | 47 |
| 2.2.3.11. | Пластиды | 49 |
| 2.2.3.12. | Клеточный центр и центриоли | 52 |
| 2.2.4. | Ядро эукариотической клетки | 53 |
| 2.2.4.1. | Ядерная оболочка | 54 |
| 2.2.4.2. | Хроматин | 55 |
| 2.2.4.3. | Ядрышко | 58 |
| 3. | Основы физиологии клетки | 68 |
| 3.1. | Клеточный метаболизм | 68 |
| 3.2. | Ферментативный характер реакций обмена | 70 |
| 3.3. | Фотосинтез | 71 |
| 3.4. | Энергетический обмен в клетке | 76 |
| 3.5. | Пластический обмен. Синтез белка | 80 |
| 3.6 | Деление клетки. митоз | 86 |
| 4. | Основы патологии клетки | 94 |
| 4.1. | Гибель клетки | 94 |
| 4.1.1. | Старение клеток | 94 |
| 4.1.2. | Некроз | 95 |
| 4.1.3. | Апоптоз | 96 |
|
| Словарь цитологических терминов | 100 |
108














Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!
Полезное для учителя

Реализация образовательных программ осуществляется с применением исключительно электронного обучения и ДОТ


