
Россия, Алтайский край, Егорьевский район, с. Сросты


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Была в сети 20.05.2024 21:10

Павлова Наталья Николаевна
учитель географии и биологии
50 лет
Местоположение
Специализация
Материал к уроку по Биологии 10-11 класс по теме "Деление клетки. Митоз"
Категория:
Биология
19.06.2022 17:17

Просмотр содержимого документа
«Материал к уроку по Биологии 10-11 класс по теме "Деление клетки. Митоз"»
Содержание работы:
Способы деления клеток – митоз.------------------ 4.
Хромосома, хроматида.------------------------------ 9.
| 1 Способы деления клеток – митоз.
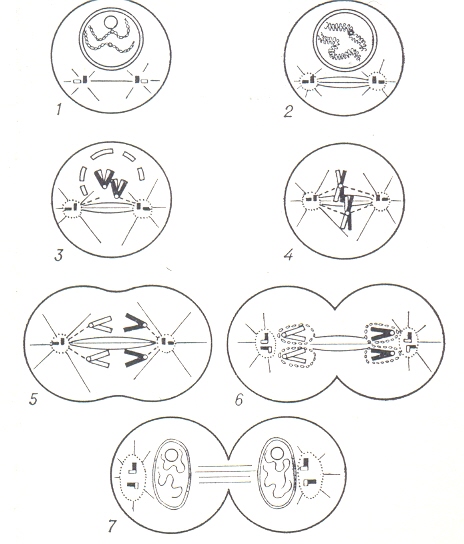
Клетка – основная структурно – функциональная единица всех живых ор - ганизмов, элементарная живая система. Она может существовать как отдель- ный организм (бактерии, простейшие, некоторые водоросли, и грибы). Размеры клетки варьируют от 0,1 – 0,25 мкм. до 155 мм. Содержимое клетки - протоплазма. В каждой клетке имеется генетический аппарат, который в клетке эукариот заключён в ядре, отделённым мембра - нами от цитоплазмы, а в клетке прокариот, лишённых оформенного ядра, в нуклеоиде. Прокариоты – организмы, клетки которых не имеют ограниченного мембраной ядра – все бактерии, включая архебактерий и цианобактерий. Эукариоты – организмы, клетки которых содержат оформленные ядра (ядерные). К эукариот относятся все высшие животные и растения, а также однокле- точные и многоклеточные водоросли, грибы и простейшие. Диаметр большинства эукариотных клеток лежит в пределах 10 – 100 мкм. Клетки эукариот способны к самовоспроизведению путём митоза; поло - вые клетки образуются путём мейоза. Мейоз – особый способ деления клеток, в результате которого происходит редукция (уменьшение) числа хромосом и переход клеток из диплоидного состояния в гаплоидное. Митоз (от греч. митос – нить. Назван так по обычно нитевидной форме хромосом.), к а р и о к и н е з, н е п р я м о е д е л е н и е к л е т к и, наиболее рас-пространённый способ воспроизведения (репродукции) клеток, обеспечиваю щий тождественное распределение генетического материала между дочерни ми клетками и преемственность хромосом в ряду клеточных поколений. Биологическое значение митоза определяется сочетанием в нём удвоения хромосом путём продольного расщепления их и равномерного распределе- ния между дочерними клетками. Началу митоза предшествует период подго-товки включающий накопление энергии, синтез дезоксирибонуклеиновой кислоты (ДНК) и репродукцию центриолей. Источником энергии служат бо-гатые энергией, или так называемые макроэргические, соединения. Митоз не сопровождается усилением дыхания, так как окислительные процессы проис ходят в интерфазе (наполнение «энергитического резервуара»). Периодичес- кое наполнение и опустошение энергетического резервуара – основа энерге- тики митоза. В 1874 году И. Д. Чистяков описал ряд стадий (фаз) митоза в спорах плау- нов, ещё не ясно представляя себе их последовательность. Детальные иссле - дования по морфологии митоза впервые были выполнены Э. Страсбургером на растениях (1876 – 1879 г.г.) и В. Флеммингом на животных (1882 г.). Единый процесс митоза подразделяют на несколько стадий, постепенно и непрерывно переходящих друг в друга: профазу, прометафазу, метафазу, анафазу, и телофазу. Длительность стадий митоза различна и зависит от ти- па ткани, физиологического состояния организма, внешних факторов. Наиболее продолжительны первая и последняя стадии. В профазе (от греч. про – до.) при исследовании под световым микроско- пом в ядре начинают выявляться хромосомы. Сначала они имеют вид клубка из перепутанных нитей. Затем нити – хромосомы укорачиваются и распола-гаются более упорядоченно, начинается процесс обособления хромосом, ко- торые становятся различными благодаря конденсации диффузного хромати- на, потере им воды. Важнейшие признаки профазы – конденсация хромосом, распад ядрышек и начало формирования веретена деления, снижение активности транскрип – ции (к концу профазы синтез РНК прекращается). Веретено деления обра – зуется либо с участием центриолей, образуя митотический аппарат (в клет – ках животных и некоторых низших растений), либо без них (в клетках выс- ших растений и некоторых простейших). У водорослей, низших, грибов и ряда, простейших веретено может формироваться внутри ядра (так назы - ваемый закрытый митоз). Прометафаза начинается распадом ядерной оболочки на фрагменты и беспорядочным движением хромосом в центральной части клетки, соответст вующей зоне бывшего ядра. При «закрытом митозе» оболочка ядра сохраня- ется в течение всего митоза. В метафазе (от греч. мета – после.) завершается формирование веретена деления. Хромосомы перестают двигаться и выстраиваются по экватору вере тена, формируя экваториальную пластинку. Синтез белка снижен на 20 – 30% по сравнению с интерфазой. На этой стадии митоза клетки наиболее чу вствительны к холоду, колхицину, его производным и другим агентам, воз- действие которых разрушает веретено деления и приводит к прекращению деления клеток (К – митоз). При низких дозах повреждающих агентов нор- мальное течение митоза восстанавливается через несколько часов после их воздействия; более высокие дозы приводят к гибели клетки, либо к её поли- плоидизации. Анафаза (от греч. ана – наверх.) - самая короткая стадия митоза. Это стадия расхождения хромосом к полюсам. Анафазное движение связано с удлинением центральных нитей веретена, раздвигающего митотические полюсы, и с укорочением хромосомальных микротрубочек митотического аппарата. Удлинение центральных нитей веретена происходит либо за счёт поляризации «запасных» макромолекул, достраивающих микротрубочки ве- ретена, либо за счёт дегидратации этой структуры. Укорочение хромосомаль ных микротрубочек обеспечивается свойствами сократительных белков ми- тотического аппарата, способных к сокращению без утолщения. Телофаза (от греч. телос – конец.) заключается в реконструкции дочер них ядер из хромосом, собравшихся у полюсов, разделении клеточного тела (цитомия, цитокинез) и окончательном разрушении митотического аппарата с образованием промежуточного тельца. Реконструкция дочерних ядер связана с деспирализацией хромосом, восстановлением ядрышка и ядерной оболочки. Цитомия осуществляется путём образования клеточной пластинки (в растительной клетке) или путём образования борозды деления (в живот- ной клетке). Механизм цитомии связан либо с сокращением желатинизиро- ванного кольца цитоплазмы, опоясывающего экватор (гипотеза «сократимо- го кольца»), либо с расширением поверхности клетки вследствие распрям- ления петлеобразных белковых цепей (гипотеза «расширения мембран»). Существенным различием телофазы от профазы является, однако, то, что каждая дочерняя хромосома состоит всего из одной хроматиды, а не из двух и поэтому содержит вдвое меньше ДНК. Достройка второй половины хро - мосом, осуществляемая путём редупликации ДНК, происходит уже в интер - фазном ядре. Продолжительность митоза зависит от размеров клеток, их плоидности, числа ядер, а также от условий окружающей среды, в часности от темпера- туры. В животных клетках митоз длится 30-60 минут, в растительных - 2-3 часа. Более длительны стадии митоза, связанные с процессами синтеза (пре- профаза, профаза, телофаза); самодвижение хромосом (метакинез, анафаза) осуществляется быстро. В результате митоза и последующей интерфазы клетки получают одинаковую наследственную информацию и содержит иде нтичные по числу, размеру и форме хромосомы с материнскими клетками. В опытах с температурно-зависимыми мутантами дрожжей и клеточных линий млекопитающих показано, что протекание митоза обуславливается активацией определённых генов и синтезом специфических РНК и белка. Иногда митоз считают только деление ядра (карионез), которое не всегда сопровождается цитотомией – образованием двух отдельных клеток. Регуляция митоза: В организме митоз контролируется системой нейро- гуморальной регуляции, которая осуществляется нервной системой, гормона ми надпочечников, гипофиза, щитовидной и половых желёз, а также местны ми факторами (продукты тканевого распада, функциональная активность клеток). Взаимодействие различных регуляторных механизмов обеспечива- ет как общие, так и местные изменения митотической активности. Митозы опухолевых клеток выходят из - под контроля нейрогуморальной регулции. Выражением регуляции митоза в связи с взаимодействием организма и сре- ды служит суточный ритм деления клеток. В большинстве органов ночных животных максимум митоза отмечается утром, а минимум – в ночное время. У дневных животных и у человека отмечается обратная динамика суточного ритма. Суточный ритм митоза – следствие цепной реакции, в которую вов- лекаются ритмические изменения внешней среды (освещённость, температу- ра , режим питания и другие), ритм функциональной активности клеток и изменения процессов обмена веществ. Нарушение митоза: при различных патологических процессах нормаль- ное течение митоза нарушается. Выделяют 3 основные вида патологии ми- тоза: 1) Повреждение хромосом ( набухание, склеивание, фрагментация, об- разование мостов, повреждение центромеров, отставание отдельных хромо- сом при движении, нарушение их спирализации и деспирализации, раннее разъединение хроматид, образование микроядер). 2) Повреждения митоти- ческого аппарата (задержка митоза в метафазе, многополюсный, моноцент- ричный и ассиметричный митоз, трёхгрупповая и полная метафазы). Особое значение в этой группе патологии митоза имеет к о л х и ц и н о в ы й митоз, или К- митоз, который называется алкалоидом колхицином (отсюда назва- ние), а также колцемидом, винбластином, винкристином, аценафтеном и дру гими так называемыми статмокинетическими ядами, используемыми в качес тве м у т а г е н о в. При К-митозе нарушаются расхождение центриолей и поляризация ими веретена деления, подвергается дезорганизации митоти -ческий аппарат, не происходит разъединения хроматид (К-пары). 3) Нару- шения цитотомии. Патологические митозы возникают после воздействия митотических ядов, токсинов, экстремальных факторов (ионизирующее из- лучение, аноксия, гипотермия), при вирусной инфекции и опухоли. Резкое увеличение числа патологического митоза типично для злокачественных опухолей. Общая схема митоза:
- Интерфаза; - Профаза; - Прометафаза; - Метафаза; - Анафаза; - Ранняя телофаза; - Поздняя телофаза; 2 Хромосома, хроматида
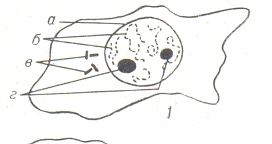
Основным морфологическим компонентом рабочего ядра является хро – матин, выявляющийся под световым микроскопом (обычно только на фик - сированных и окрашенных препаратах) в виде сети из тонких длинных ни - тей, а также мелких гранул. Своё название он получил за способность свя - завать многие красители. Хроматин (от греч. chroma – цвет, краска) представляет собой нуклеопро - теидные нити, из которых состоят хромосомы клеток эукариот; во время де- ления клетки формируется в определённые видимые структуры в хромосо- мах. Термин хроматин был введён немецким гистологом в 1888 году В. Флем - мингом. В современной цитологии под хроматином подразумевают хромо - сомное вещество ядра клетки в интерфазе (между последовательными её де- лениями), так как хромосомы в этот период клеточного цикла под микроско- пом плохо обнаруживаются. В состав хроматина в определённых пропорци ях входят: дезоксирибонуклеиновая кислота ( ДНК) - (30 – 45%), рибонукле иновая кислота (РНК), гистоны (30 – 50%), и негистоновые белки (4 – 33%). Основной структурный компонент хроматина – дезоксирибонуклепротеид- ные нити (ДНП) диаметром 100- 200 А, основу каждой из которых, по мне- нию большинства исследователей, составляет одна молекула ДНК. Предло- жено две модели тонкой структуры элементарной нити хроматина: супеспи- ральная (американские учёные Д. Пардон, М. Уилкинс, 1972 г.) и глобуляр- ная (американские учёные А. Корнберг, А. Л. Олинс и Д. Э. Олинс, 1974г.) На электронных микрофотографиях хроматин напоминает бусы, «снизан - ные» из нуклеосом – частиц диаметром около 10 нм. Высшие порядки струк- турной организации хроматина (хромосомы) образуются из линейного пучка элементарных нитей хроматина – нуклеосом – за счёт суперспирализации образования петель прикрепления к «осевому скелету» из негистоновых бел- ков. В этих процессах участвуют гистоны, ионы металлов и так далее. Разли- чие между активным и неактивным хроматином связывают, прежде всего, с различиями состава и со структурными переходами последнего (главным об- разом плотностью упаковки). Возможно, что эти типы различаются нуклео- сомной организацией. Различают генетически активный хроматин (эухроматин) и неактивный (гетерохроматин). В ядрах клеток особей женского пола многих организ – мов (в частности, млекопитающих животных и человека) обнаружены круп- ные плотные глыбки хроматина, которых нет у особей мужского пола. Такой хроматин назван «половым хроматином». Половой хроматин – участки хроматина, определяющие различие интерфаз-ных ядер у особей разных полов, связанные с особенностями структуры или функционирования половых хромосом. Различают Y – половой хроматин (Y – хроматин) и X – половой хроматин (X – хроматин). Y – хроматин - структурный гетерохроматин Y - хромосомы человека, выявляющийся в интерфазном ядре с помощью флюоорохромов в ультрофиолетовом свете. X-хроматин или тельце Барра, - интенсивно красящаяся основными краси- телями структура (0,7 – 1,2 мкм.), которая находится в ядрах разных типов клеток самок, образован в норме одной из двух половых хромосом гомога -метного пола. Эта хромосома спирализирована и вследствии этого не актив- на. При наличии большего числа X- хромосом такой инактивации подверга- ются все, кроме одной X-хромосомы. Поэтому количество телец полового хроматина на единицу меньше X-хромосом и служит диагностическим при- знаком при определении их количества. Подобный механизм образования полового хроматина у большинства млекопитающих. Хромосомы (от хромо… и сома), органоиды клеточного ядра, являющиеся носителями генов и определяющие наследственные свойства клеток и орга-низмов. Полный набор хромосом в клетке, характерный для данного организ ма, называют к а р и о т и п о м. В любой клетке тела большинства живот- ных и растений каждая хромосома представлена дважды: одна из них полу- чена от отца, другая- от матери при слиянии ядер половых клеток в процессе оплодотворения. Такие хромосомы называют гомологичными, а набор гомо- логичных хромосом – диплоидным. В хромосомном наборе клеток раздель- нополых организмов присутствует пара (или несколько пар) половых хромо- сом, как правило, различающихся у разных полов по морфологическим при- знакам; остальные хромосомы называют а у т о с о м а м и . У млекопитающих в половых хромосомах локализованы гены, определяю- щие пол организма; у плодовой мушки дрозофилы пол определяется соотно- шением половых хромосом и аутосом (балансовая теория определения пола) Хромосомы способны к самовоспроизведению, обладают структурной и фун кциональной индивидуальностью и сохраняют её в ряду поколений. Первоначально хромосомы были описаны как интенсивно окрашивающиеся основными красителями плотные тельца (немецкий учёный В. Вальдейер 1888 год). Однако оказалось, что внешний вид хромосом существенно меняется на разных стадиях клеточного цикла, и как компактные образования с характер ной морфологией, хромосомы чётко различимы в световом микроскопе лишь в период клеточного деления – в метафазе митоза и мейоза. Основу хромосом на всех стадиях клеточного цикла составляют хромонемы - нитевидные структуры, которые во время деления клетки плотно закруче- ны, обуславливая спирализацию хромосом, а в неделящейся клетке раскруче-ны (деспирализованы). Половые клетки, образовавшиеся в результате мейоза, содержат только одну из двух гомологичных хромосом. Число хромосом сильно варьирует: от двух до нескольких сотен хромосом и составляют хромосомные наборы разных видов. Каждый вид организмов обладает характерным и постоянным набором хромосом в клетке, закреплённым в эволюции данного вида, а его изменения происходят только в результате мутаций. При завершении деления клетки, разошедшиеся к её полюсам, хромосомы разрыхляются и окружаются ядерной мембраной. В период между двумя делениями клетки (эта стадия клеточного цикла называется и н т е р ф а з о й) деспирализация хромосом продолжается и они становятся малодоступными для наблюдения в световой микроскоп. Морфология хромосом эукариот существенно отличается от таковой у прокариот и вирусов. Прокариоты (доядерные) и вирусы содержат обычно одну линейную и кольцевую хромосому, которая не имеет надмолекулярной укладки и не отделена от цитоплазмы ядерной оболочкой. Понятие хромосомы к генетическому аппарату прокариот применимо лишь условно, так как оно сформировалось при изучении хромосом эукариот и подразумевает наличие в хромосомах не только сложного комплекса биополимеров (нуклеиновых кислот и белков), но и спе цифической надмолекулярной структуры. Изменения внешнего вида хромо- сом в клеточном и жизненном циклах обусловлены особенностями функцио- нирования хромосом. Общий же принцип их организации, индивидуальность и непрерывность хромосом в ряду клеточных поколений и организмов сохраняются неизменными. Доказательства тому получены при биохимичес ких, цитологических и генетических исследованиях хромосом разных орга- низмов. Они легли в основу хромосомной теории наследственности. Молекулярные основы строения хромосом: Значение хромосом как кле- точных органоидов, ответственных за хранение, воспроизведение и реализа- цию наследственной информации, определяется свойствами биополимеров, входящих в их состав. Первая молекулярная модель хромосом была предло- жена в 1928 году Н. К.Кольцовым, предугадавшим принципы их организа- ции. Запись наследственной информации в хромосомах обеспечивается стро ением молекулы дезоксирибонуклеиновой кислоты (ДНК), её генетическим кодом. В хромосомах сосредоточено около 99% всей ДНК клетки, остальная часть ДНК находится в других клеточных органоидах, определяя цитоплаз- матическую наследственность. ДНК в хромосомах эукариот находится в ком плексе с основными белками - гистонами и с негистоновыми белками, ко- торые обеспечивают сложную упаковку ДНК в хромосоме и регуляцию её способности к синтезу рибонуклеиновых кислот (РНК) – транскрипции. Хромосома в интерфазе: Хромосома выполняет свои основные функции - репродукцию и транскрипцию - в интерфазе, поэтому строение хромосомы на этой стадии клеточного цикла представляет особый интерес. В интерфазе хромосомы хорошо различимы потому, что в связи с активным синтезом РНК многие участки хромосомы (т. н. эухроматин) сильно раскручены; дру-гие же (гетерохроматин) не участвуют в синтезе РНК и продолжают сохра- нять плотную упаковку. В эухроматиновыхучастках, помимо элементарных дезоксирибонуклепротеидных нитей (ДНП), имеются рибонуклепротеидные частицы диаметром 200- 500 А, называются РНП – гранулами, интерграну- лами перихроматиновыми гранулами. Эти частицы представляют собой фор му упаковки РНК, синтезированной на хромосоме и соединённой с белком, и служат для завершения образования информационной РНК и переноса её в цитоплазму. Для изучения интерфазных хромосом используют биохимичес- кие методы выделения вещества хромосом – хроматина и разделения его на эухроматин и гетерохроматин. Гетерохроматин (от гетеро… и хроматин), участки хроматина, находящие в конденсированном (плотно упакованном) состоянии в течение всего клеточ-ного цикла. Интенсивно окрашиваются ядерными красителями и хорошо ви- дны в световой микроскоп даже во время интерфазы.) сестринских хрома - тид. Гетерохроматические районы хромосом, как правило, реплицируются позже эухроматиновых и не транскрибируются, то есть генетически весьма инертны. Различают факультативный и конститутивный (структурный) гетерохматин. Факультативный гетерохроматин – присутствует только в одной из гомологич-ных хромосом. Пример гетерохроматина такого типа – вторая X-хромосома у женских особей млекопитающих, которая в ходе раннего эмбриогенеза инактивируется вследствие её необратимой конденсации. Структурный гетерохроматин – содержится в обоих гомологичных хромосо – мах, локализирован преимущественно в экспонированных участках хромо – сомы – в центромере, теломере, ядрышковом организаторе (во время интер- фазы он располагается неподалёку от ядерной оболочки), обеднён генами, обогащён сателлитной ДНК и может инактивировать расположенные по со – седству гены (так называемый эффект положения). Этот тип гетерохромати - на очень вариабелен как в пределах одного вида, так и в пределах несколь - ких видов. Он может влиять на синопсис хромосом, частоту индуцирован - ных разрывов и рекомбинацию. Участкам структурного гетерохроматина свойственна адгезия (слипание). Эухроматин (от греч. eu – хорошо, полностью и хроматин), участки хромосом, сохраняющие деспирализованное состояние в покоящемся ядре (в интерфа – зе) и специализирующиеся при делении клеток (в профазе); содержит боль- шинство генов и потенциально способных к транскрипции. Эухроматин от- личается от гетерохроматина меньшим содержанием метилированных осно- ваний и блоков повторяющихся последовательностей ДНК, большим коли- чеством негистоновых белков и ацетилированных молекул гистонов, менее плотной упаковкой хромосомного материала, что, как полагают, особенно важно для активности эухроматина и делает его потенциально более доступ- ным для ферментов, обеспечивающих транскрипцию. Эухроматин может приобретать свойства факультативного гетерохроматина – инактивировать- ся, что является одним из способов регуляции генной активности.. В кариотипе кроме половых хромосом, аутосом, различают ядрышкообра- зующие хромосомы; у некоторых видов могут существовать добавочные хро мосомы, число которых непостоянно и которые не содержат генов, свойст- венных данному виду. А также для изучения интерфазных хромосом используют электронно-мик- роскопическое исследование интактных ядер и изолированного хроматина; как модели интерфазных хромосом используют гигантские хромосомы типа ламповых щёток из ооцистов животных и многонитчатые (политенные) хромосомы двукрылых. (В процессе развития многоклеточных организмов хромосо- мы могут приобретать своеобразную форму и в некоторых случаях имеют специальное название, например политенные хромосомы типа ламповых щёток и др.) Политения – (от поли… и лат. taenia – повязка, лента), образование в ядре соматичес ких клеток некоторых двукрылых, простейших и растений гигантских многонитча – тых (политенных) хромосом, превышающих по размерам в сотни раз обычные. За счёт многократной репликации исходной хромосомы без последующего её расхожде – ния число хромонем (иногда свыше 1000) и количество ДНК увеличивается, что и при-водит к увеличению диаметра и длины хромосом. Политения впервые описана Э. Баль-биани в 1881 году. Благодаря неравномерной спирализации в политенных хромосомах образуются диски (тёмные поперечные полосы), выявляемые при окраске хромосом. Число, размер и характер расположения дисков специфичны для вида. Политению используют для построения карт хромосом, обнаружения хромосомных перестроек; сравнение цитологических карт политенных хромосом позволяет определять видовую принадлежность особей разных популяций и способствует пониманию процессов микроэволюции и видообразования.
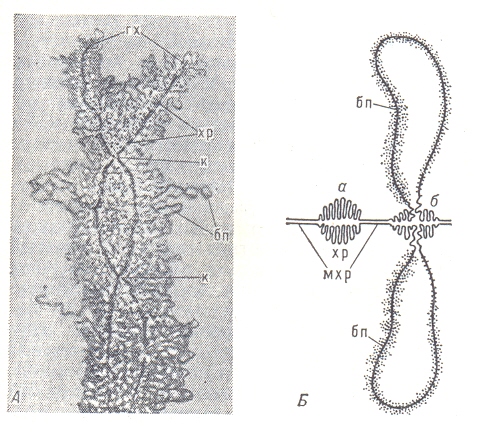
А – Структура хромосом типа лампо- вых щёток (из женских половых клеток ток тритона) в профазе мейоза; гх-гомо - логичные хромосомы, ещё сохраняющие в отдельных местах коньюгацию (к); хр – хромомеры; бп – боковые петли хро момер (где происходит синтез РНК).
Б – неактивная (а) и функционирую- щая (б) хромомеры; последняя образует боковые петли (бп); мхр – межхромер- ные участки хромосомы.
В хромосомах типа ламповых щёток неактивные участки имеют вид плот- но упакованных структур – хромомер (см. рис) , которые обнаруживаются и в хромосоме соматических клеток, особенно в профазе митоза, и рассматри- ваются как характерные морфологические, а возможно и функциональные, единицы хромосом. В участках хромосом, активно синтезирующих РНК, хромомеры раскручиваются и образуют боковые петли, в которых молеку- лы РНК, соединяясь с белком, образуют рибонуклеопротеиды (РНП) – час- тицы, представляют собой форму упаковки генных продуктов и различаю- щиеся в отдельных боковых петлях по размерам и морфологическим призна- кам. В политенных хромосомах, возникающих в тканях двукрылых и некото рых растений за счёт многократной репликации (удвоения) исходной хромо- сомы без последующего расхождения дочерних хромосом, неактивные учас- тки имеют форму дисков, а активные образуют вздутия – пуфы. В пуфах, так же как и в хромосомах типа ламповых щёток, содержатся частицы РНП диа- метром 200-500А. Изучение политенных хромосом в разных тканях и на разных стадиях раз- вития двукрылых показало, что число и набор активных пуфов имеют ткане- вую и видовую специфичность. Это значит, что хотя все клетки многоклето- чного организма имеют одинаковый набор генов, линейно расположенных в каждой хромосоме, набор активных и неактивных в синтезе РНК участков хромосомы различается в каждом типе клеток и на разных стадиях развития, т. е. один и тот же участок находится в одних тканях в эухроматическом, в других – в гетерохроматическом состоянии.
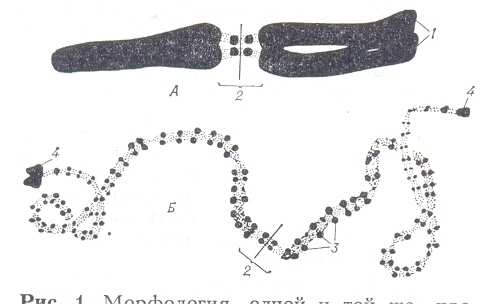
Морфология одной и той же хромосомы в метафазе митоза (А) и профазе мейоза (Б); 1 – хроматиды; 2 – центромера; 3 – хромомеры; 4 – теломеры (крупные хромо меры на концах хромосомы).
Утолщённые, плотно спирализированные участки дезоксирибонуклеопро- теидных нитей хромосомы, образующиеся в результате местного скручива –ния, называются хромомерами ( от хромо и греч. meros – часть). Они интенсив но окрашиваются ядерными красителями. Под микроскопом хорошо разли – чимы в профазе мейоза, особенно в пахитене, имеют вид тёмноокрашенных гранул, расположенных, подобно бусинам, по длине хромосом. Хромомер – ный рисунок (расстояние между хромомерами и их размер) используется для идентификации хромосом. У некоторых видов растений обнаружены очень крупные хромомеры (состоят из гетерохроматина и называются «узел ками», или вздутиями), которые служат хромосомными маркерами (метка – ми) при цитогенических исследованиях. Генетическое же значение хромоме-ров и межхромоммерных районов остаётся неясным. Половые хромосомы – хромосомы, определяющие различие кариотипов особей разных полов у раздельнополых организмов. Пол, имеющий 2 одина-ковые половые хромосомы, обозначаемые обычно как X-хромосомы, назы - вают гомогаметным. Гетерогаметный пол у разных видов животных и расте-ний имеет либо одну X-хромосому (тип XO), либо пару различающихся по - ловых хромосом – X и Y (тип XY). Как в типе XY (человек, другие млекопи-тающие, дрозофила), как и в типе XO (клопы, кузнечики) в большинстве случаев гетерогаметен мужской пол. В этом случае у самок в результате мейоза образуются гаметы, содержащие всего по одной X-хромосоме, у самцов одни гаметы формируются с X-, другие с Y-хромосомой или без по - ловых хромосом. Оплодотворение яйцеклетки сперматозоидом, несущим X-хромосому, приводит к образованию XX-зиготы, из которой развивается женская особь; оплодотворение сперматозоидом, не содержащим X-хромо – мы, приводит к появлению мужской особи. У птиц, бабочек, некоторых пресмыкающихся и земноводных гомогаметен мужской пол, а гетерогаметен женский. Половые хромосомы содержат гены, определяющие не только половые, но и другие признаки организма, которые называют сцепленными с полом. Y-хромосома (посравнению с X-хромосомой) часто обеднена гена -нами, содержит много структурного гетерохроматина и, как правило, мень -ше по размеру. Большинство генов X-хромосомы не представлены в Y-хро –мосоме, но доза их обычно компенсируется у гомогаметного пола. Нерасхождение половых хромосом у одного из родителей в момент образо - вания половых клеток приводит к нарушению развития организма. В результате нарушения расхождения хромосом в митозе или мейозе под действием высокой или низкой температуры, ионизирующих излучений, химических веществ возникает полиплоидия (от греч. polyploos-многократный и eidos-вид) – наследственное изменение, заключающееся в кратном увеличе – нии числа наборов хромосом в клетках организмов. Наиболее часто встреча- етя у растений простейших, а из многоклеточных животных – у дождевых червей. При полиплоидии наблюдается отклонение от диплоидного числа хромосом в соматических клетках и от гаплоидного – в половых; могут воз - никать клетки, в которых каждая хромосома представлена трижды (3n – три-плоиды), четырежды (4n – тетраплоиды), пять раз (5n – пентаплоиды) и так далее. Различают автополиплодию (кратное увеличение числа наборов хро - мосом одного вида), характерную, как правило, для видов с вегетативным способом размножения (автополиплоиды стерильны в связи с нарушением конъюгации гомологичных хромосом в процессе мейоза), и аллополиплои –дию (изменение числа наборов хромосом на основе межвидовой гибридиза – ции), при которой обычно происходит удвоение числа хромосом у бесплоид-ного диплоидного гибрида, и он становится в результате этого плодовитым. Хромосомные перестройки, хромосомные мутации, структурные измене- ния хромосом, возникающие вследствие разрывов в них, сопровождающих- ся в большинстве случаев воссоединением образовавшихся кусков в иных сочетаниях, чем в исходных хромосомах. Структурные перестройки могут происходить как в пределах одной хромосомы, так и между гомологичными и негомологичными хромосомами состоят в выпадении (делеция), или удво- ении (дупликация)какого либо участка хромосомы, в перенесении его в дру- гой сегмент хромосомы (транслокация); наконец участок может оказаться перевёрнутым на 180 градусов, оставаясь в той же хромосоме (инверсия). Делеции и дуплекации нарушают генный баланс, что ведёт к изменению признаков организма. Инверсии, изменяющие лишь порядок изменения ген- ов в хромосоме, и транслокации, при которых гены перемещённого участка попадают в другую группу сцепления, не нарушают генного баланса и не из-меняют фенотипических признаков организма. При мейозе у гетерозигот, содержащих одну нормальную хромосому и одну с инверсией, сближение этих хромосом затруднено, кроссинговер между ними подавлен или идёт с пониженной частотой, нередко возникают анеуплоидные гаметы, поэтому такие организмы отличаются пониженной плодовитостью по сравнению с гомозиготами, у которых обе хромосомы данной пары несут инверсию или обе нормальны. Гетерозиготы по хромосомам, несущим транслокацию, дают много анеуплоидных гамет, поэтому плодовитость их ниже, чем у гомозиго- тных организмов. Хромосомные перестройки могут возникать спонтанно, но частота их резко возрастает под влиянием физических и химических факторов, и играют боль шую роль в эволюции организмов: дуплекации представляют главный источ ник увеличения числа генов; инверсии и транслокации могут вести к генети- ческой изоляции гомозиготных по ним особей, более плодовитых, чем гете- розиготы. При всех хромосомных перестройках иногда наблюдается эффект положения гена, заключающийся в том, что ген, перенесённый в новое мес- то хромосомы, изменяет своё действие на фенотип организма. Хромосомные перестройки могут быть использованы в практических целых для изменения групп сцепления генов, определяющих хозяйственно ценные признаков орга низмов.
Хромосомный набор, совокупность хромосом, заключённая в ядре любой клетки тела растительного и животного организма; характеризуется постоян ным для каждого биологического вида числом хромосом, определённой их величиной и морфологическими особенностями. Гетерохроматиновый участок хромосомы, сохраняющий в интерфазном ядре клетки спирализованную структуру хромонемы называется хромоцент ром или кариосомой. Под микроскопом при окрашивании ядерными краси - телями имеет вид глыбки. В зависимости от положения гетерохроматина на хромосоме (теломерное или центромерное) хромоцентры занимают соответ - ственно приближённое или удалённое расположение по отношению к эквато ру клетки во время деления, сохраняя это положение до следующего деле – ния. Хромоцентр может быть образован одним гетерохроматиновым райо – ном или несколькими районами. Количество хромоцентра меняется в зави – симости от количества хромосом в ядре, а также от стадии клеточного цикла Набор кариосом отражает количество неактивных в синтезе РНК участков хромосом и соответственно особенности функционирования ядер разных типов клеток. Функции хромоцентра неясны. Структурным элементом хромосомы является хроматида (от греч. chro - matos – цвет, краска), формирующийся в интерфазе ядра клетки в результа - те удвоения хромосом. Наиболее хорошо различима во время метафазы ми - тоза , когда хромосома состоит из двух хроматид; после деления центроме - ры хроматиды расходятся в дочерние ядра и становятся самостоятельными хромосомами. В мейозе гомологичные хромосомы, сближаясь попарно, обра зуют структуру из четырёх хроматид (тетраду). Различия между хроматида -ми могут возникать в результате кроссинговера (несестринские обмены) или реципрокного обмена участками хроматиды одной хромосомы (сестринс - кие обмены). Частота таких обменов повышается под действием некоторых физических, химических и физиологических факторов. Согласно однонитчатой модели хромосомы, каждая хроматида содержит в поперечнике одну суперспирализованную и конденсированную двуцепотча- тую молекулу дезоксирибонуклеиновой кислоты (ДНК); многонитчатая мо- дель хромосомы предполагает наличие в поперечнике каждой хроматиды несколько молекул ДНК (в этом случае различают полухроматиды, четверть хроматиды и т.д.). Экспериментально более подтверждена однонитчатая мо- дель. Хромосомная теория наследственности: Это теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и пред -ставляют собой материальную основу наследственности, то есть преемствен ность свойств организмов в ряду поколений определяется преемственностью хромосом. Хромосомная теория наследственности возникла в начале 20 века на основе клеточной теории и использования для изучения наследственных свойств организмов гидрологического анализа. В 1902 году У. Сентон в США, обративший внимание на параллелизм в по- ведении хромосом и менделевских так называемых «наследственных факто- ров»,и Т. Бовери в Германии выдвенули хромосомную теорию наследствен- ности, согласно которой менделеевские наследственные фарторы (назван- ные впоследствии генами) локализованы в хромосомах. С развитием этой теории было установлено, что гены, расположенные в одной хромосоме, составляют одну группу сцепления и должны наследова-ться совместно; число групп сцепления равно числу пар хромосом, постоян- ному для каждого вида организмов; признаки, зависящие от сцепленных ге- нов, также наследуются совместно. Согласно представлениям школы Моргана, гены являются дискретными далее неделимыми носителями наследственной информации. Однако откры- тие в 1925 году советскими учёными Г. А. Надсоном и Г. С. Филипповым, а в 1927 году американским учёным Г.Меллером влияния рентгеновских лу - чей на возникновения наследственных изменений (мутаций) у дрозофилы, а также применение рентгеновских лучей для ускорения мутационного процес са у дрозофилы позволили советским учёным А. С. Серебровскому, Н. П. Ду бинину и др. сформулировать в 1928- 30 гг. представление о делимости гена на более мелкие единицы, расположенные в линейной последовательности и способные к мутационным изменениям. Хромосомная теория наследственности. Объясняя закономерности наследо- вания признаков у животных и растительных организмов, играет важную роль в сельскохозяйственной науке и практике. Она вооружает селекционе- ров методами выведения пород животных и сортов растений с заданными свойствами. Некоторые положения хромосомной теории наследственности позволяют более рационально вести с/х производство. Так, явление сцеплен- ного с полом наследования ряда признаков у с/х животных позволило до изобретения методов искусственного регулирования пола у тутового шелко- пряда выбраковывать коконы менее продуктивного пола. До разработки способа разделения цыплят по полу исследования клоаки – отбраковывать петушков и т. п. Важнейшее значение для повышения урожайности многих с/х культур имеет использование полиплоидии. На знании закономерностей хромосомных перестроек основывается изучение наследственных заболева- ний человека. Хромосомные болезни, наследственные заболевания, обусловленные изме нением числа или структуры хромосом. Частота хромосомных болезней среди новорождённых детей около 1%. Многие изменения хромосом несов- местимы с жизнью и являются частой причиной спонтанных абортов и мерт- ворождений. При спонтанных абортах обнаружено около 20% эмбрионов с аномальными кариотипами. Изменение числа хромосом происходит в резу -льтате нерасхождения их в мейозе или при делении клеток на ранней стадии развития оплодотворённого яйца. Хромосомные болезни могут быть связаны с излишком генетического мате- риала (полисомия- наличие одной или несколько добавочных хромосом; по- липлоидия; дупликация); с утраченной части генетического материала (нул -лисомия, моносомия, делеция); с хромосомными перестройками (транслока- ция; различные перестановки участков хромосом). Различают также группы хромосомных болезней, обусловленных изменениями половых и неполовых хромосом. Наиболее распространённые аномалии первой группы у женщин – синдром Шерешевского-Тернера (моносомияХ) (возникает задержка полово- го развития, бесплодие) и синдром трисомии Х (некоторое снижение интеллекта, расстройства менструального цикла); у мужчин – синдром Клайнфельтера, характеризующий наличием лишней Х- хромосомы. Частота аномалий по половым хромосомам у мертворождённых детей составляет 2,7%, что в 25 раз выше, чем среди новорождённых. Среди аутосомных аномалий с нарушением числа хромосом выделяют три- сомные синдромы: синдром трисомии хромосом группы D (13-15-е пары), или синдром Патау; синдром трисомии группы Е (18-я пара) – Эдвардса, и Дауна болезнь (трисомия по 21 хромосоме). Указанные хромосомные болез -ни проявляются различными уродствами; задержкой физического и умствен- ного развития; пороками развития умственных органов. Подобные больные живут, как правило, недолго, погибают от вторичных инфекций. Тяжесть клинической картины при синдромах, вызванные структурными изменения-ми хромосом,как правило, коррелирует с количеством избыточного или не- достающего хромосомного материала. Специфика патологических проявле- ний зависит от того, какая хромосома вовлечена в процесс перестройки. Наряду с типичными хромосомными болезнями описано большое количест- во (около 200) синдромов, вызванных сложными типами хромосомных аббе раций. Единственно надёжный метод диагностики хромосомных болезней – цито- генетическое исследование кариотипа, а при изменении числа половых хро- мосом – дополнительное исследование полового хроматина.
| |
|
|
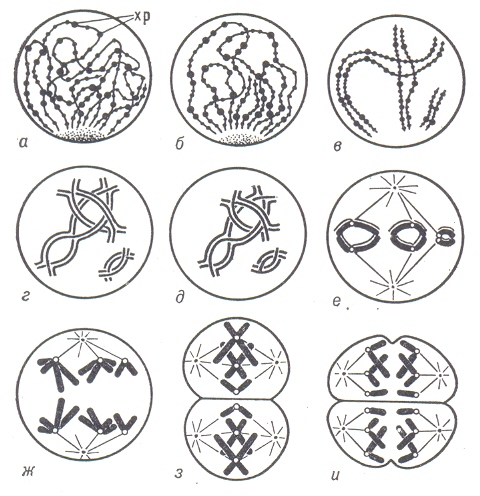
Схема мейоза.
Этот тип деления клетки характеризуется длительной стадией профазы (а - д). При подготовке к метафазе (г, д) гомологические хромосомы начинают отталкиваться, а затем быстро следуют два мейотических деления (е - и); хр – хромомеры.
|









Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!

