
Россия, Калининград


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Была в сети 06.06.2018 00:05

Мудрицкая Светлана Викторовна
заместитель директора, учитель биологии
78 лет
Местоположение
Специализация
Учебное пособие "Мир клетки" Модуль 3 "Основные принципы строения эукариотической клетки" к Программе "Путешествие в мир клетки"
Категория:
Биология
28.05.2018 21:25
Учебник:
Биология. Общая биология. Профильный уровень. 10 класс. Захаров В.Б. и др. М.: 2010. - 352 с.

Просмотр содержимого документа
«Учебное пособие "Мир клетки" Модуль 3 "Основные принципы строения эукариотической клетки" к Программе "Путешествие в мир клетки"»
2.2. Общий план строения эукариотической клетки
Клетки многоклеточного животного организма разнообразны. Значительно различаются по форме, размерам, по строению и функциям клетки крови, мышечные и нервные клетки, сперматозоиды и клетки желез.
Клетки растений также достаточно разнообразны: это клетки эпидермы и мезофила, ситовидные трубочки и волокна колленхимы.
О
днако исходя из постулата Р. Вирхова о гомологичности клеток, можно говорить о едином плане строения всех клеток: они состоят из поверхностного аппарата, ядра и цитоплазмы.
эукариотическая клетка
| поверхностный аппарат клетки
| Внутреннее содержимое клетки (протопласт) | |||||
| Цитоплазматическая мембрана | Ядро
| Цитоплазма | ||||
| Ядерная оболочка | Нуклеоплазма | Гиалоплазма | Органеллы | Включения | ||
| Кортикальный слой цитоплазмы
Гликокаликс (животные клетки) или
Клеточная стенка (растительные клетки) |
Хроматин Ядерный матрикс Ядрышко
|
Мембраные |
Немембранные |
Трофические Секреторные Экскреторные Пигментные | ||
| ЭПС – гладкая и гранулярная Лизосомы Митохондрии Пластиды Комплекс Гольджи Глиоксисомы Синаптические пузырьки | Рибосомы Центриоли
| |||||
Все эукариотические клетки отличаются значительным развитием мембранных структур.
2.2.1. Морфофункциональные особенности клеточных мембран
Клеточные мембраны (6-10 нм) - важнейший компонент живого содержимого клетки, выполняют барьерную, транспортную и рецепторную функции.
Все без исключения клеточные мембраны построены по общему принципу: это тонкие липопротеидные пленки, состоящие из двойного слоя липидных молекул, в который включены молекулы белка. На долю липидов (жироподобных веществ) приходится 20-80%, на долю белков 20-80%. В состав многих мембран входят углеводы, количество которых достигает 2-10%. К липидам относится большая группа органических веществ, обладающих плохой растворимостью в воде (гидрофобность) и хорошей растворимостью в органических растворителях (липофильность). Характерными представителями липидов, встречающихся в клеточных мембранах, являются фосфолипиды (глицерофосфатиды), гликолипиды, сфингомиелины и холестерин. В растительных клетках холестерин не обнаружен, его там заменяют фитостерины (эргостерол). Характерной особенностью липидов мембран является разделение их молекулы на неполярные хвосты, состоящие из жирных кислот, и заряженные полярные головки, образованные многоатомными спиртами. На поверхности воды растворы полярных липидов, растекаясь, образуют мономолекулярную пленку, в которой в водную фазу будут направлены заряженные (гидрофильные) головки, а неполярные хвосты будут обращены в сравнительно гидрофобную воздушную фазу. Смеси разных липидов формируют бимолекулярные слои или мембраны толщиной около 3,5 нм, где периферические зоны слоя, смотрящие в воду, будут содержать полярные головки, а незаряженные хвосты будут образовывать общую гидрофобную центральную часть образовавшейся мембраны. Таким образом, способность липидов самопроизвольно образовывать мембранные структуры определяется их химической структурой. Как искусственные, так и естественные мембраны не представляют собой плоские слои, они всегда замкнуты сами на себя и образуют полые вакуоли, пузырьки, плоские замкнутые мешки или трубчатые образования. Липидные мембраны служат непроницаемым барьером для любых заряженных молекул, даже для ионов солей. Это определяет основное функциональное свойство мембран - служить преградой для свободной диффузии через слой липидов. Это свойство может быть использовано для практических целей. Так при смешивании липидов в водной среде образуется масса полых мембранных пузырьков - липосом. Жидкость, попавшая внутрь этих пузырьков, уже не может свободно обмениваться с жидкостью, находящейся снаружи. Таким образом, искусственные мембраны липосом можно «загрузить» лекарственными веществами, которые могут в нужных концентрациях поступать к клеткам.
Белки – второй компонент клеточных мембран. Количество белков в разных мембранах может быть различным. Так в мембранах митохондрий доля белков составляет около 75%, а в плазматической мембране клеток миелиновой оболочки - около 25%. Белковые молекулы как бы вкраплены в билипидный слой мембраны. Часть из них связана с липидными головками с помощью ионных связей и поэтому легко экстрагируется из мембран растворами солей. Легко экстрагируемые белки расположены на поверхности мембран и их называют периферийными. Большая часть белков взаимодействует с липидами в составе мембран на основе гидрофобных связей. Оказалось, что многие мембранные белки состоят как бы из двух частей: из участков, богатых полярными аминокислотами, и участков, обогащенных неполярными аминокислотами. Такие белки в липидных слоях мембран располагаются так, что их неполярные участки погружены в «жирную» часть мембраны, где находятся гидрофобные участки липидов. Полярная часть таких белков взаимодействует с головками липидов и обращена в сторону водной фазы, поэтому такие белки, связанные с липидами путем гидрофобных взаимодействий, практически не экстрагируются в водных фазах. Эти белки мембран называют интегральными. Размер интегральных мембранных белков в среднем равен 8 нм, но встречаются крупные белки - до 35 нм величиной (белок тилакоидов хлоропластов). Интересно, что большая часть липидных молекул (70%) не связана с белками, так что белковые молекулы как бы плавают в «липидном озере». Липидные молекулы также двигаются вдоль липидного слоя, могут вращаться вокруг своей оси. Гарт Л. Николс и С. Дж. Синджер в 1972 г. предложили жидкостно-мозаичную модель строения элементарной мембраны.
Н
есмотря на поразительную схожесть строения различных мембран, построенных по принципу липидного бислоя с вмонтированными в него белками, физические и химические свойства разных мембран различны. Это связано с тем, что в разных мембранах общий состав липидов значительно различается, что определяет особые свойства мембран.
Разные мембраны клетки могут отличаться друг от друга по количеству липидов. Так, плазматическая мембрана содержит 35-40% липидов, а мембраны митохондрий - 27-29%. Самое высокое содержание липидов в плазматической мембране шванновских клеток, образующих миелиновую оболочку нервов, - до 80%. Было обнаружено, что клеточные мембраны сильно отличаются друг от друга по составу липидов. Так, плазматические мембраны клеток животных богаты холестерином (до 30%), в то время как мембраны митохондрий, наоборот, богаты фосфолипидами и бедны холестерином.
Особенно отличаются мембраны по составу белков, которые, главным образом, определяют функциональные свойства мембран. По биологической роли мембранные белки можно разделить на группы: ферменты, рецепторные белки, структурные и транспортные белки.
Набор ферментов в составе мембран может быть очень велик и разнообразен (например, в плазматической мембране клеток печени обнаружено не менее 24 различных ферментов). В разных мембранах существует характерный набор ферментов. Например, в плазматической мембране, как и во многих других, локализуется K+-Na+-зависимая АТФаза, участвующая в транспорте ионов. В митохондриях специфическим является набор белков - переносчиков электронов и фермент АТФ-синтетаза, обеспечивающие окислительное фосфорилирование и синтез АТФ.
Рецепторные белки специфически связываются с теми или иными веществами и как бы их «узнают». Это белки-рецепторы для гормонов, для узнавания поверхности соседних клеток, вирусов, фагов у бактерий и т.д. К этой группе относятся фоторецепторные белки. Рецепторные белки входят в состав любых мембран.
Со стороны цитоплазмы мембраны связаны через примембранные или собственно мембранные интегральные белки с разнообразными белковыми структурами цитоплазмы. К ним относятся в первую очередь компоненты цитоскелета. Это позволяет не только сделать мембраны более жесткими, но и обеспечивает подвижность мембран, создавая возможности для их транспортных функций. Во многих эпителиальных клетках специальные белки плазматической мембраны связываются с элементами цитоскелета и участвуют в образовании целого ряда межклеточных соединений (десмосомы, адгезивный контакт и др.).
2.2.2. Поверхностный аппарат клетки
Плазмолемма среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, ограничивающая клетку снаружи. Плазматическая мембрана исполняет роль барьера между внутриклеточным содержимым и внешней средой; она ограничивает свободный поток низко- и высокомолекулярных веществ в обе стороны через мембрану. Плазмолемма - это структура «узнающая» различные химические вещества и регулирующая избирательно транспорт этих веществ в клетку и из нее. Плазмолемма участвует в делении клетки, в образовании ресничек и микроворсинок, а так же в формировании межклеточных контактов.
Г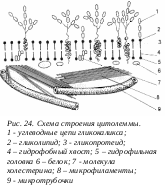 ликокаликс представляет собой внешний по отношению к липопротеидной мембране слой, содержащий полисахаридные цепочки мембранных интегральных белков - гликопротеидов. Эти цепочки содержат такие углеводы как манноза, глюкоза и др. Углеводные гетерополимеры образуют ветвящиеся цепочки, между которыми могут располагаться выделенные из клетки гликолипиды и протеогликаны. Слой гликокаликса сильно обводнен, имеет желеподобную консистенцию, что значительно снижает в этой зоне скорость диффузии различных веществ. Здесь же могут «застревать» выделенные клеткой гидролитические ферменты, участвующие во внеклеточном расщеплении полимеров (внеклеточное пищеварение) до мономерных молекул, которые затем транспортируются в цитоплазму через плазматическую мембрану.
ликокаликс представляет собой внешний по отношению к липопротеидной мембране слой, содержащий полисахаридные цепочки мембранных интегральных белков - гликопротеидов. Эти цепочки содержат такие углеводы как манноза, глюкоза и др. Углеводные гетерополимеры образуют ветвящиеся цепочки, между которыми могут располагаться выделенные из клетки гликолипиды и протеогликаны. Слой гликокаликса сильно обводнен, имеет желеподобную консистенцию, что значительно снижает в этой зоне скорость диффузии различных веществ. Здесь же могут «застревать» выделенные клеткой гидролитические ферменты, участвующие во внеклеточном расщеплении полимеров (внеклеточное пищеварение) до мономерных молекул, которые затем транспортируются в цитоплазму через плазматическую мембрану.
В электронном микроскопе гликокаликс имеет вид рыхлого волокнистого слоя, толщиной 3-4 нм, покрывающего всю поверхность клетки. Он обнаружен практически у всех животных клеток, но степень его выраженности различна. Особенно хорошо гликокаликс развит в щеточной каемке клеток всасывающего эпителия кишечника.
Механическая устойчивость плазматической мембраны, кроме того, обеспечивается структурой примыкающего к ней со стороны цитоплазмы кортикального слоя и внутриклеточных фибриллярных структур. Кортикальный (от слова cortex - кора, кожица) слой цитоплазмы, лежащий в тесном контакте с плазмолеммой, имеет ряд особенностей. Здесь, при толщине его в 0,1-0,5 мкм, отсутствуют рибосомы и мембранные пузырьки, но в большом количестве встречаются фибриллярные элементы цитоплазмы - микрофиламенты и, часто, микротрубочки. Основным фибриллярным компонентом кортикального слоя является сеть микрофибрилл актиновых белков и вспомогательных белков, необходимых для движения участков цитоплазмы.
Транспорт ионов через плазмалемму проходит за счет участия в этом процессе мембранных транспортных белков - пермеаз. Эти белки могут вести транспорт в одном направлении одного вещества (унипорт), или нескольких веществ одновременно (симпорт), или же вместе с импортом одного вещества выводить из клетки другое (антипорт). Так, например, глюкоза может входить в клетки симпортно вместе с ионом Na+.
Т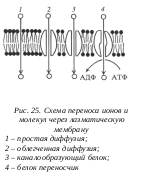 ранспорт ионов может происходить по градиенту концентрации – пассивно, без дополнительной затраты энергии. Так, например, в клетку проникает ион Na+ из внешней среды, где его концентрация выше, чем в цитоплазме. В случае пассивного транспорта некоторые мембранные транспортные белки образуют молекулярные комплексы, каналы, через которые растворенные молекулы проходят через мембрану за счет простой диффузии по градиенту концентрации. Часть этих каналов открыта постоянно, а другая часть может закрываться или открываться в ответ либо на связывание с сигнальными молекулами, либо на изменение внутриклеточной концентрации ионов. В других случаях специальные мембранные белки-переносчики избирательно связываются с тем или иным ионом и переносят его через мембрану - это облегченная диффузия.
ранспорт ионов может происходить по градиенту концентрации – пассивно, без дополнительной затраты энергии. Так, например, в клетку проникает ион Na+ из внешней среды, где его концентрация выше, чем в цитоплазме. В случае пассивного транспорта некоторые мембранные транспортные белки образуют молекулярные комплексы, каналы, через которые растворенные молекулы проходят через мембрану за счет простой диффузии по градиенту концентрации. Часть этих каналов открыта постоянно, а другая часть может закрываться или открываться в ответ либо на связывание с сигнальными молекулами, либо на изменение внутриклеточной концентрации ионов. В других случаях специальные мембранные белки-переносчики избирательно связываются с тем или иным ионом и переносят его через мембрану - это облегченная диффузия.
В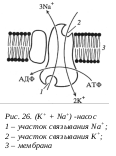 клетках существуют мембранные белковые переносчики, которые работают против градиента концентрации, затрачивая при этом энергию АТФ. Такой тип работы носит название активного транспорта, и он осуществляется с помощью белковых ионных насосов. В плазматической мембране находится двухсубъединичная молекула (K+ + Na+)-насоса, которая одновременно является и АТФазой. Этот насос при работе откачивает за один цикл 3 иона Na+ и закачивает в клетку 2 иона K+ против градиента концентрации. При этом затрачивается одна молекула АТФ, идущая на фосфорилирование АТФазы, в результате чего Na+ переносится через мембрану из клетки, а K+ получает возможность связаться с белковой молекулой и затем переносится в клетку.
клетках существуют мембранные белковые переносчики, которые работают против градиента концентрации, затрачивая при этом энергию АТФ. Такой тип работы носит название активного транспорта, и он осуществляется с помощью белковых ионных насосов. В плазматической мембране находится двухсубъединичная молекула (K+ + Na+)-насоса, которая одновременно является и АТФазой. Этот насос при работе откачивает за один цикл 3 иона Na+ и закачивает в клетку 2 иона K+ против градиента концентрации. При этом затрачивается одна молекула АТФ, идущая на фосфорилирование АТФазы, в результате чего Na+ переносится через мембрану из клетки, а K+ получает возможность связаться с белковой молекулой и затем переносится в клетку.
Такая постоянная работа пермеаз и насосов создает в клетке постоянную концентрацию ионов и низкомолекулярных веществ. В сочетании с активным транспортом ионов через плазматическую мембрану происходит транспорт различных сахаров, нуклеотидов и аминокислот.
Макромолекулы, такие как белки, нуклеиновые кислоты, полисахариды, липопротеидные комплексы и другие, сквозь клеточные мембраны не проходят. В клетку же или из одного мембранного компартмента в другой макромолекулы попадают заключенными внутри вакуолей или везикул. Такой везикулярный перенос можно разделить на два вида: экзоцитоз - вынос из клетки макромолекулярных продуктов и эндоцитоз - поглощение клеткой макромолекул.
При эндоцитозе определенный участок плазмалеммы захватывает, как бы обволакивает, внеклеточный материал, заключает его в мембранную вакуоль, возникшую за счет впячивания плазматической мембраны. В такую первичную вакуоль могут попадать любые биополимеры, части клеток или даже целые клетки, где затем и распадаются до мономеров. Основное биологическое значение эндоцитоза - это получение строительных блоков за счет внутриклеточного переваривания, которое осуществляется после слияния первичной эндосомы с лизосомой.
Эндоцитоз формально разделяют на пиноцитоз и фагоцитоз. Фагоцитоз - захват и поглощение клеткой крупных частиц (иногда даже клеток или их частей) - был впервые описан И.И. Мечниковым. Фагоцитоз, способность захватывать клеткой крупные частицы, встречается среди клеток животных как одноклеточных (например, амебы, некоторые хищные инфузории), так и для специализированных клеток многоклеточных животных - фагоцитов. Пиноцитоз вначале определялся как поглощение клеткой воды или водных растворов разных веществ. Сейчас известно, что как фагоцитоз так и пиноцитоз протекают очень сходно, и поэтому употребление этих терминов может отражать лишь различия в объемах, массе поглощенных веществ. Общее для этих процессов то, что поглощенные вещества на поверхности плазматической мембраны окружаются мембраной в виде вакуоли - эндосомы, которая перемещается внутрь клетки.
Дальнейшая судьба эндосом может быть различной: часть из них может возвращаться к поверхности клетки и сливаться с ней, но большая часть вступает в процесс внутриклеточного пищеварения. Первичные эндосомы содержат в основном захваченные в жидкой среде чужеродные молекулы и не содержат гидролитических ферментов. Эндосомы могут сливаться друг с другом, при этом увеличиваясь в размере. Они затем сливаются с первичными лизосомами, которые вводят в полость эндосом ферменты, расщепляющие (гидролизующие) различные биополимеры.
Фагоцитоз является вариантом эндоцитоза и связан с поглощением клеткой крупных агрегатов макромолекул вплоть до живых или мертвых клеток. При фагоцитозе происходит образование больших эндоцитозных вакуолей - фагосом, которые затем, сливаясь с лизосомами, образуют фаголизосомы.
Плазматическая мембрана принимает участие в выведении веществ из клетки с помощью экзоцитоза - процесса, обратного эндоцитозу. В случае экзоцитоза, внутриклеточные продукты, заключенные в вакуоли или пузырьки и отграниченные от гиалоплазмы мембраной, подходят к плазматической мембране. В местах их контактов плазматическая мембрана и мембрана вакуоли сливаются, и пузырек опустошается в окружающую среду. С экзоцитозом связано выделение синтезированных в клетке разнообразных веществ. Клетки могут вырабатывать и выделять во внешнюю среду низкомолекулярные соединения (ацетилхолин, биогенные амины и др.), а также макромолекулы: пептиды, белки, липопротеиды, пептидогликаны и др.. Выделенные из клеток путем экзоцитоза гидролитические ферменты могут сорбироваться в слое гликокаликса и обеспечивать примембранное внеклеточное расщепление различных биополимеров и органических молекул. Огромное значение примембранное неклеточное пищеварение имеет для животных. Было обнаружено, что в кишечном эпителии млекопитающих в зоне щеточной каемки всасывающего эпителия, особенно богатой гликокаликсом, обнаруживается огромное количество разнообразных ферментов.
2.2.3. Цитоплазма
Цитоплазма - это тот компонент клетки, который остается, если исключить ядро. Цитоплазма клетки многокомпонентна. Даже в световой микроскоп в цитоплазме живой клетки видны вкрапления, неоднородности, частички, но особенно неоднородность цитоплазмы видна при изучении ее в электронном микроскопе.
Формально структуру цитоплазмы подразделяют на три части: органеллы, включения, гиалоплазма (основная плазма или цитозоль).
О рганеллы - обязательные для любой клетки компоненты, без которых клетка просто не может поддерживать свое существование, они имеют постоянную структуру и выполняют определенные функции. Включения – это необязательные компоненты, которые представляют собой или отложения запасных веществ (гликоген, желточные гранулы) или скопление продуктов метаболизма (пигменты, кристаллы солей и др. в растительных клетках).
рганеллы - обязательные для любой клетки компоненты, без которых клетка просто не может поддерживать свое существование, они имеют постоянную структуру и выполняют определенные функции. Включения – это необязательные компоненты, которые представляют собой или отложения запасных веществ (гликоген, желточные гранулы) или скопление продуктов метаболизма (пигменты, кристаллы солей и др. в растительных клетках).
Органеллы и включения погружены в гиалоплазму - жидкую фазу цитоплазмы клетки.
2.2.3.1. ГиалоплазмаТермины гиалоплазма (от hyaline - прозрачный), основная плазма, матрикс цитоплазмы или цитозоль, обозначают очень важную часть клетки, ее истинную внутреннюю среду. Гиалоплазма - не просто разбавленный водный раствор органических соединений: его состав весьма сложен, а консистенция приближается к гелю (желе). Гели - это структурированные коллоидные системы с жидкой дисперсной средой. Частицы дисперсной фазы соединены между собой в рыхлую пространственную сетку, которая содержит в своих ячейках дисперсную среду, лишая текучести систему в целом. Гель гиалоплазмы под воздействием внешних условий (температура, давление) или внутренних факторов может переходить в менее вязкую, более жидкую фазу - в золь или раствор. Такие гель-золь переходы очень характерны для гиалоплазмы. Отдельные зоны гиалоплазмы могут менять свое агрегатное состояние в зависимости от условий или функции. Отдельные молекулы белков-тубулинов могут быть диспергированы в гиалоплазме, но в определенные моменты они начинают собираться и строить длинные трубчатые структуры - микротрубочки. Этот процесс самосборки микротрубочек обратим: при изменении условий последние распадаются до мономерных молекул тубулинов. Таким же образом в гиалоплазме могут возникать и распадаться различные фибриллярные, нитчатые комплексы белковых молекул.
Функциональное значение гиалоплазмы очень велико. Здесь находятся ферменты, участвующие в синтезе аминокислот, нуклеотидов, жирных кислот, сахаров. В гиалоплазме происходит синтез и отложение запасного полисахарида гликогена, накопление запасных жировых капель. Здесь же происходят процессы гликолиза и синтез части АТФ.
В гиалоплазме на рибосомах и полирибосомах, не связанных с мембранами ЭПС, происходит синтез белков, которые транспортируются в различные участки клетки. Здесь осуществляется синтез всех белков клеточного ядра, большей частью белков митохондрий и пластид. Кроме структурных белков и ферментов в цитозоле в растворенном состоянии содержится огромное количество аминокислот, нуклеотидов и других строительных блоков биополимеров, а также множество метаболитов - промежуточных продуктов, возникающих при синтезе и распаде макромолекул. Гиалоплазма содержит большое количество ионов неорганических соединений, таких как Na+, K+, Ca2+, Cl-, HCO3-, HPO42- и др. При этом концентрация этих ионов строго регулируется мембранными компонентами клетки.
Формально, по морфологическим признакам, органеллы можно разделить на две группы: мембранные и немембранные. Мембранные органеллы так же представлены двумя вариантами: одномембранные и двумембранные. К первым относятся органеллы вакуолярной системы – эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы и другие специализированные вакуоли. К двумембранным органеллам относятся митохондрии и пластиды. К немембранным органеллам принадлежат рибосомы, клеточный центр клеток низших растений и животных. Клеточный скелет или цитоскелет - постоянный компонент клетки, его выраженность может значительно меняться в течение клеточного цикла.
2.2.3.2. Эндоплазматическая сеть
Эндоплазматическая сеть – ЭПС (ретиклум ЭР) - сквозная сеть двухконтурных мембран, соединенных друг с другом, образующая каналы, вакуоли, мешочки и трубочки. Эта внутриклеточная мембранная структура была открыта в 1945 году К. Портером. В 50-х гг., при использовании метода ультратонких срезов удалось выяснить структуру эндоплазматического ретикулюма и обнаружить его неоднородность. Подобный электронно-микроскопический анализ позволил выделить два типа ЭПС: гранулярная (шероховатая) и агранулярная или гладкая ЭПС.
Г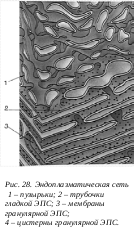 ранулярная ЭПС представлена замкнутыми мембранами, которые образуют на сечениях вытянутые мешки, цистерны или же имеют вид узких (около 20 нм) каналов. Отличительной особенностью гранулярной ЭПС, является то, что ее мембраны со стороны гиалоплазмы покрыты мелкими (около 20 нм) темными, почти округлыми частицами, гранулами - рибосомами. На мембранах рибосомы расположены в виде полисом (множество рибосом, объединенных одной информационной РНК). Это синтезирующие белок рибосомы прикрепляются к мембранам большой субъединицей. Гранулярная ЭПС может в клетках быть представлена или в виде редких разрозненных мембран или же в виде скоплений - это эргастоплазма. Первый тип гранулярноой ЭПС характерен для малодифференцированных клеток или клеток с низкой метаболической активностью. Эргастоплазма характерна для клеток, активно синтезирующих белки. В клетках печени каналы гранулярной ЭПС собраны в отдельные зоны - тельца Берга, а в нервных клетках формирует тигроид или вещество Ниссля. Количество рибосом на ЭПС четко связано с ее синтетической активностью. Например, на мембранах ЭПС в клетке несекретирующей молочной железы связывается до 25% клеточных рибосом, в лактирующей железе их количество возрастает до 70%. Уменьшение числа рибосом наблюдается и при различных патологических состояниях клеток (при алкогольном хроническом отравлении происходит уменьшение числа связанных рибосом на 25%). Рибосомы, связанные с мембранами ЭПС, участвуют в синтезе белков, выводимых из данной клетки, т.е. «экспортируемых» белков.
ранулярная ЭПС представлена замкнутыми мембранами, которые образуют на сечениях вытянутые мешки, цистерны или же имеют вид узких (около 20 нм) каналов. Отличительной особенностью гранулярной ЭПС, является то, что ее мембраны со стороны гиалоплазмы покрыты мелкими (около 20 нм) темными, почти округлыми частицами, гранулами - рибосомами. На мембранах рибосомы расположены в виде полисом (множество рибосом, объединенных одной информационной РНК). Это синтезирующие белок рибосомы прикрепляются к мембранам большой субъединицей. Гранулярная ЭПС может в клетках быть представлена или в виде редких разрозненных мембран или же в виде скоплений - это эргастоплазма. Первый тип гранулярноой ЭПС характерен для малодифференцированных клеток или клеток с низкой метаболической активностью. Эргастоплазма характерна для клеток, активно синтезирующих белки. В клетках печени каналы гранулярной ЭПС собраны в отдельные зоны - тельца Берга, а в нервных клетках формирует тигроид или вещество Ниссля. Количество рибосом на ЭПС четко связано с ее синтетической активностью. Например, на мембранах ЭПС в клетке несекретирующей молочной железы связывается до 25% клеточных рибосом, в лактирующей железе их количество возрастает до 70%. Уменьшение числа рибосом наблюдается и при различных патологических состояниях клеток (при алкогольном хроническом отравлении происходит уменьшение числа связанных рибосом на 25%). Рибосомы, связанные с мембранами ЭПС, участвуют в синтезе белков, выводимых из данной клетки, т.е. «экспортируемых» белков.
Роль гранулярной ЭПС заключается не только в участии в синтезе белков на рибосомах его мембран, но и в процессе обособления этих синтезированных белков, в их изоляции от других белков клетки. Эта функциональная особенность гранулярной ЭПС обеспечивает выделение синтезированных белков с помощью вакуолей комплекса Гольджи. В гранулярной ЭПС п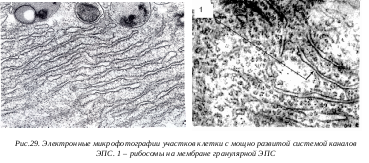 роисходит синтез белков, которые, встраиваясь в ее мембрану, становятся интегральными мембранными белками.
роисходит синтез белков, которые, встраиваясь в ее мембрану, становятся интегральными мембранными белками.
Сегодня можно сказать, что важнейшей функцией гранулярной ЭПС, вне зависимости от специализации или тканевой принадлежности клеток, является функция образования, построения клеточных мембран, которая заключается в том, что элементы гранулярной ЭПС синтезируют все мембранные белки, синтезируют липидный компонент мембран, но, кроме того, именно здесь происходит сборка липопротеидных мембран. От того, какие интегральные и периферические белки будут синтезироваться на рибосомах ЭПС, и от того, какие фосфолипиды будут синтезироваться и включаться в мембрану, будет зависеть тип образующегося нового участка мембраны: будет ли он компонентом гладкой ЭПС, мембран комплекса Гольджи, лизосомы. или плазматической мембраны.
Участки гранулярной ЭПС, которые расположены в зоне около комплекса Гольджи (КГ), теряют рибосомы и образуют мембранные выступы, от которых отпочковываются мелкие вакуоли, содержащие синтезированные в ЭПС белки. Отделившиеся от ЭПС вакуоли теряют белковую оболочку, сливаются друг с другом и транспортируются с помощью микротрубочек к цис-зоне комплекса Гольджи, где и сливаются с его мембранами.
2.2.3.3. Рибосомы
Рибосома представляет собой элементарную клеточную машину синтеза любых белков клетки. В отличие от других органелл цитоплазмы (пластид, митохондрий, клеточного центра, мембранной вакуолярной системы и др.), рибосомы в клетке многочисленны: за клеточный цикл их образуется 1х107 штук. Рибосомы - это сложные рибонуклеопротеидные частицы, в состав которых входит множество молекул индивидуальных белков и несколько молекул РНК. Р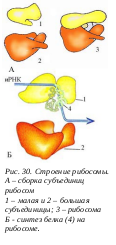 ибосомы прокариот и эукариот по своим размерам отличаются, хотя и обладают общими принципами организации и функционирования. Полная, работающая рибосома, состоит из двух неравных субъединиц: большой и малой. Размер полной прокариотической рибосомы составляет 20 х 17 х 17 нм, эукариотической - 25 х 20 х 20. Полная прокариотическая рибосома имеет коэффициент седиментации 70S и диссоциирует на две субъединицы: 50S и 30S. Полная эукариотическая рибосома, 80S рибосома, диссоциирует на 60S и 40S субъединицы. Малая рибосомная субъединица имеет палочковидную форму с несколькими небольшими выступами. Большая субъединица похожа на полусферу с тремя торчащими выступами. При образовании полной рибосомы, малая субчастица ложится одним концом на один из выступов большой субъединицы, а другим в ее желобок. В состав малых субъединиц входит по одной молекуле РНК, а в состав большой – несколько: у прокариот – две, а у эукариот – 3 молекулы.
ибосомы прокариот и эукариот по своим размерам отличаются, хотя и обладают общими принципами организации и функционирования. Полная, работающая рибосома, состоит из двух неравных субъединиц: большой и малой. Размер полной прокариотической рибосомы составляет 20 х 17 х 17 нм, эукариотической - 25 х 20 х 20. Полная прокариотическая рибосома имеет коэффициент седиментации 70S и диссоциирует на две субъединицы: 50S и 30S. Полная эукариотическая рибосома, 80S рибосома, диссоциирует на 60S и 40S субъединицы. Малая рибосомная субъединица имеет палочковидную форму с несколькими небольшими выступами. Большая субъединица похожа на полусферу с тремя торчащими выступами. При образовании полной рибосомы, малая субчастица ложится одним концом на один из выступов большой субъединицы, а другим в ее желобок. В состав малых субъединиц входит по одной молекуле РНК, а в состав большой – несколько: у прокариот – две, а у эукариот – 3 молекулы.
Рибосомные РНК обладают сложной вторичной и третичной структурой, образуя сложные петли и шпильки на комплементарных участках, что приводит к самоупаковке, самоорганизации этих молекул в сложное по форме тело.
2.2.3.4. Комплекс Гольджи (КГ)
В 1898 году итальянский ученый Камилло Гольджи, используя свойства связывания тяжелых металлов (осмия и серебра) с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал «внутренним сетчатым аппаратом». Обычно элементы комплекса Гольджи расположены около ядра, вблизи клеточного центра. Участки комплеса Гольджи, имеют в некоторых клетках вид сложных сетей, где ячейки связаны друг с другом или представлены в виде отдельных темных участков, лежащих независимо друг от друга, имеющих вид палочек, зерен, вогнутых дисков. В электронном микроскопе видно, что комплекс Гольджи представлен мембранными структурами, собранными вместе в небольшой зоне. Отдельная зона скопления этих мембран называется диктиосомой. В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены в виде стопки плоские мембранные мешки, или цистерны, между которыми располагаются тонкие прослойки гиалоплазмы. Каждая отдельная цистерна имеет диаметр около 1 мкм; в центре ее мембраны могут быть сближены (25 нм), а на периферии имеют расширения, ампулы. Количество таких мешков в стопке обычно не превышает 5-10. Кроме плотно расположенных плоских цистерн в зоне комплекса Гольджи наблюдается множество вакуолей. Мелкие вакуоли встречаются главным образом в периферических участках; иногда видно, как они отшнуровываются от ампулярных расширений на краях плоских цистерн.
1898 году итальянский ученый Камилло Гольджи, используя свойства связывания тяжелых металлов (осмия и серебра) с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал «внутренним сетчатым аппаратом». Обычно элементы комплекса Гольджи расположены около ядра, вблизи клеточного центра. Участки комплеса Гольджи, имеют в некоторых клетках вид сложных сетей, где ячейки связаны друг с другом или представлены в виде отдельных темных участков, лежащих независимо друг от друга, имеющих вид палочек, зерен, вогнутых дисков. В электронном микроскопе видно, что комплекс Гольджи представлен мембранными структурами, собранными вместе в небольшой зоне. Отдельная зона скопления этих мембран называется диктиосомой. В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены в виде стопки плоские мембранные мешки, или цистерны, между которыми располагаются тонкие прослойки гиалоплазмы. Каждая отдельная цистерна имеет диаметр около 1 мкм; в центре ее мембраны могут быть сближены (25 нм), а на периферии имеют расширения, ампулы. Количество таких мешков в стопке обычно не превышает 5-10. Кроме плотно расположенных плоских цистерн в зоне комплекса Гольджи наблюдается множество вакуолей. Мелкие вакуоли встречаются главным образом в периферических участках; иногда видно, как они отшнуровываются от ампулярных расширений на краях плоских цистерн.
Принято различать в зоне диктиосомы проксимальный или формирующийся, цис-участок, и дистальный или зрелый, транс-участок. Между ними располагается средний или промежуточный участок комплекса Гольджи.
В секретирующих клетках обычно комплекс Гольджи поляризован: его проксимальная часть обращена к цитоплазме и ядру, а дистальная - к поверхности клетки.
К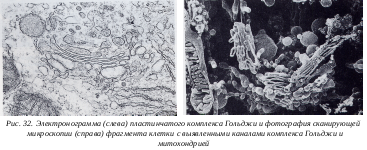 омплекс Гольджи является промежуточным звеном между собственно синтезом секретируемого белка и выведением его из клетки. Синтезированный на рибосомах экспортируемый белок отделяется и накапливается внутри цистерн ЭПС, по которым он транспортируется к зоне мембран комплекса Гольджи. Здесь от гладких участков ЭПС отщепляются мелкие вакуоли, содержащие синтезированный белок, которые поступают в зону вакуолей в проксимальной части диктиосомы. В этом месте вакуоли могут сливаться друг с другом и с плоскими цис-цистернами диктиосомы. Происходит перенесение белкового продукта уже внутри полостей цистерн КГ. По мере созревания белков в цистернах комплекса Гольджи, они с помощью мелких вакуолей переносятся от цистерн к цистерне в дистальную часть диктиосомы, пока не достигают трубчатой мембранной сети в транс-участке диктиосомы. В этом участке происходит отщепление мелких пузырьков, содержащих уже зрелый продукт. Отделившиеся мелкие пузырьки сливаются друг с другом, образуя секреторные вакуоли. После этого секреторные вакуоли начинают двигаться к поверхности клетки, соприкасаются с плазматической мембраной, с которой сливаются их мембраны, и, таким образом, содержимое этих вакуолей оказывается за пределами клетки. Этот процесс напоминает пиноцитоз, только с обратной последовательностью стадий, и называется экзоцитоз.
омплекс Гольджи является промежуточным звеном между собственно синтезом секретируемого белка и выведением его из клетки. Синтезированный на рибосомах экспортируемый белок отделяется и накапливается внутри цистерн ЭПС, по которым он транспортируется к зоне мембран комплекса Гольджи. Здесь от гладких участков ЭПС отщепляются мелкие вакуоли, содержащие синтезированный белок, которые поступают в зону вакуолей в проксимальной части диктиосомы. В этом месте вакуоли могут сливаться друг с другом и с плоскими цис-цистернами диктиосомы. Происходит перенесение белкового продукта уже внутри полостей цистерн КГ. По мере созревания белков в цистернах комплекса Гольджи, они с помощью мелких вакуолей переносятся от цистерн к цистерне в дистальную часть диктиосомы, пока не достигают трубчатой мембранной сети в транс-участке диктиосомы. В этом участке происходит отщепление мелких пузырьков, содержащих уже зрелый продукт. Отделившиеся мелкие пузырьки сливаются друг с другом, образуя секреторные вакуоли. После этого секреторные вакуоли начинают двигаться к поверхности клетки, соприкасаются с плазматической мембраной, с которой сливаются их мембраны, и, таким образом, содержимое этих вакуолей оказывается за пределами клетки. Этот процесс напоминает пиноцитоз, только с обратной последовательностью стадий, и называется экзоцитоз.
В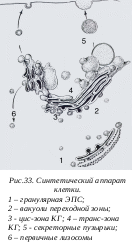 комплексе Гольджи происходит не просто «перекачка» продуктов из одной полости в другую, но и постепенно идет их «созревание», модификация белков, которая заканчивается «сортировкой» продуктов, направляющихся или к лизосомам, или к плазматической мембране, или к секреторным вакуолям.
комплексе Гольджи происходит не просто «перекачка» продуктов из одной полости в другую, но и постепенно идет их «созревание», модификация белков, которая заканчивается «сортировкой» продуктов, направляющихся или к лизосомам, или к плазматической мембране, или к секреторным вакуолям.
2.2.3.5. Лизосомы
Лизосомы не представляют собой в клетках самостоятельных структур, они образуются за счет активности ЭПС и комплекса Гольджи, они участвуют в процессах внутриклеточного расщепления биологических макромолекул.
Лизосомы были открыты Де Дюв в 1955году. Лизосомы представляют собой окруженные липопротеидной мембраной пузырьки, содержащие около 40 гидролитических ферментов: протеиназы, нуклеазы, гликозидазы, фосфорилазы, фосфатазы, сульфитазы и др. Под электронным микроскопом видно, что лизосомы представлены пузырьками (0,2-0,4 мкм), ограниченными одиночной мембраной, с очень разнородным содержимым внутри. Во фракции лизосом встречаются пузырьки с гомогенным, бесструктурным содержимым, и пузырьки, заполненные плотным веществом, содержащим в свою очередь вакуоли; часто можно видеть внутри лизосом не только участки мембран, но и фрагменты митохондрий и ЭПС.
Было обнаружено, что среди различных по морфологии лизосом можно выделить четыре типа: первичные лизосомы, вторичные лизосомы, аутофагосомы и остаточные тельца. Первичные лизосомы представляют собой мелкие мембранные пузырьки размером около 100 нм, заполненные бесструктурным веществом, содержащим набор гидролаз и в том числе кислую фосфатазу, - характерный для лизосом фермент. Эти мелкие вакуоли, первичные лизосомы, трудно отличить от мелких вакуолей на периферии зоны комплекса Гольджи. С помощью ряда точных экспериментов установили, что в дальнейшем первичные лизосомы сливаются с фагоцитарными или пиноцитозными вакуолями, эндосомами, образуя вторичную лизосому или внутриклеточную пищеварительную вакуоль. При этом содержимое первичной лизосомы сливается с полостью эндоцитозной вакуоли, и гидролазы первичной лизосомы получают доступ к веществам, которые они и начинают расщеплять.
Разнообразие по величине и по структуре клеточных лизосом связано в первую очередь с разнообразием вторичных лизосом - продуктов слияния эндоцитозных вакуолей с первичными лизосомами. Таким образом, вторичные лизосомы представляют собой не что иное, как внутриклеточные пищеварительные вакуоли, ферменты которых доставлены с помощью мелких первичных лизосом. Поэтому от типа поглощенных веществ или частичек зависит размер и внутренняя структура таких лизосом.
Р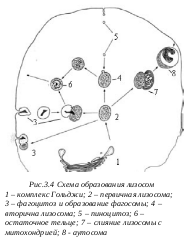 асщепление, переваривание биогенных макромолекул внутри лизосом может идти в ряде клеток не до конца. В этом случае в полостях лизосом происходит накопление непереваренных продуктов, происходит переход вторичных лизосом в телолизосомы, или остаточные тельца. Часто в остаточных тельцах наблюдается вторичная структуризация непереваренных липидов, которые образуют сложные слоистые структуры. Там же происходит отложение пигментных веществ. У человека при старении организма в клетках мозга, печени и в мышечных волокнах в телолизосомах происходит отложение «пигменте старения» - липофусцина.
асщепление, переваривание биогенных макромолекул внутри лизосом может идти в ряде клеток не до конца. В этом случае в полостях лизосом происходит накопление непереваренных продуктов, происходит переход вторичных лизосом в телолизосомы, или остаточные тельца. Часто в остаточных тельцах наблюдается вторичная структуризация непереваренных липидов, которые образуют сложные слоистые структуры. Там же происходит отложение пигментных веществ. У человека при старении организма в клетках мозга, печени и в мышечных волокнах в телолизосомах происходит отложение «пигменте старения» - липофусцина.
Аутолизосомы (аутофагосомы) постоянно встречаются в клетках простейших, растений и животных. По своей морфологии их относят к вторичным лизосомам, но с тем отличием, что в составе этих вакуолей встречаются фрагменты или даже целые цитоплазматические структуры, такие, как митохондрии, пластиды, элементы ЭПС, рибосомы, гранулы гликогена и т.д. Процесс образования аутофагосом еще недостаточно ясен. По одним представлениям, первичные лизосомы могут выстраиваться вокруг клеточной органеллы, сливаться друг с другом и таким образом отделять ее от соседних участков цитоплазмы - участок оказывается отделенным мембраной и заключенным внутри такой сложной лизосомы. Процесс аутофагоцитоза связан с отбором и уничтожением измененных, «сломанных» клеточных компонентов. В этом случае лизосомы выполняют роль внутриклеточных чистильщиков, контролирующих дефектные структуры. Значительно возрастает число аутофагосом при различных повреждениях клеток.
2.2.3.6. Агланулярная (гладкая) эндоплазматическая сеть
Агранулярная ЭПС представляет собой часть мембранной вакуолярной системы клетки, представлена мембранами, образующими мелкие вакуоли и трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярной, на мембранах гладкой ЭПС нет рибосом. Диаметр вакуолей и канальцев гладкой ЭПС обычно около 50-100 нм. Выраженность мембран гладкой ЭПС может быть неодинаковой как для различных клеток, так и внутри одной клетки. В клетках эпителия кишечника гладкая ЭПС локализуется главным образом в апикальной, верхней части клетки, вблизи всасывающей поверхности. В клетках печени зоны гладкой ЭПС часто связаны с местами отложения гликогена.
Неоднократно была установлена непрерывность перехода между гладкой ЭПС и гранулярной. Цистерны гранулярной ЭПС теряют на своей поверхности рибосомы и становятся гладкими. При этом такой участок цистерны делается неровным, начинает ветвиться, переходя в трубочки и канальцы агранулярной ЭПС. Гладкая ЭПС является вторичной по отношению к гранулярной и происходит из последней.
Деятельность гладкой ЭПС связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов.
Агранулярная эндоплазматическая сеть особенно развита в клетках, секретирующих стероиды, например, в клетках коркового вещества надпочечника, интерстициальных клетках семенников, участвующих в синтезе стероидных гормонов, а также в клетках сальных желез.
В печени часто увеличение зон гладкой ЭПС связано с рядом патологических процессов в клетках. Так, при барбитуратных отравлениях, при действии различных канцерогенов или ядовитых веществ, при действии больших доз гормональных препаратов клетки печени теряют характерную для них базофилию цитоплазмы, в них падает содержание РНК и появляются в цитоплазме оксифильные зоны. В электронном микроскопе эти зоны представлены скоплениями мембран гладкой ЭПС. Это явление связано с тем, что в этих местах происходят процессы разрушения различных вредных веществ.
В поперечнополосатых мышцах каналы гладкой ЭПС окружают каждую миофибриллу. Здесь ЭПС выполняет специальную функцию депонирования ионов кальция.
Среди высших растений агранулярная эндоплазматическая сеть встречается в клетках тканей, участвующих в синтезе и транспорте терпенов, стероидов, липидов.
45
Комплекты видеоуроков для учителей








Скачать
© 2018, Мудрицкая Светлана Викторовна 1051 10
Похожие файлы
Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!

