
УЗБЕКИСТАН, ДЖИЗАК


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до 10.06.2025
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Была в сети 20.04.2023 08:06

Мавлонова Сунажон Хужаязовна
Старший Преподователь по биологии
58 лет
Местоположение
Специализация
Взаимодействие аллельных генов. взаимодействие неаллельных генов.
Категория:
Биология
06.09.2020 14:32

Просмотр содержимого документа
«Взаимодействие аллельных генов. взаимодействие неаллельных генов.»
ВЗАИМОДЕЙСТВИЕ АЛЛЕЛЬНЫХ ГЕНОВ
 Согласно закономерностям, установленным Менделем, гены способны к стабильному воспроизведению и фенотипическому проявлению. Гены проявляют свое действие независимо от других генов. Могут происходить мутации генов и возникать различные рекомбинации. Между геном и признаком - сложная связь. Действие генов специфично. Один ген может отвечать за один признак.
Согласно закономерностям, установленным Менделем, гены способны к стабильному воспроизведению и фенотипическому проявлению. Гены проявляют свое действие независимо от других генов. Могут происходить мутации генов и возникать различные рекомбинации. Между геном и признаком - сложная связь. Действие генов специфично. Один ген может отвечать за один признак.
Продуктом функции генов является белок - фермент, катализирующий определенную биохимическую реакцию, играющую важную роль в формировании признака в определенных условиях среды.
-Один ген может отвечать за несколько признаков, проявляя плейотропное действие. Например, синдром Марфана. Это наследственное заболевание, развивающееся от присутствия в генотипе одного измененного гена. Заболевание проявляется следующими признаками: подвывихом хрусталика глаза, аневризмой аорты, изменением длины пальцев - появлением "паучьих пальцев", высоким сводом стопы. В основе лежит нарушение развития соединительной ткани.
Выраженность плейотропного действия гена зависит от биохимической реакции, которую катализирует фермент, синтезируемый под контролем данного гена.
Проявление признаков есть результат взаимодействия различных биохимических реакций. Эти взаимодействия могут быть связаны с аллельными и неаллельными генами.
Взаимодействие аллельных генов может происходить по типу:
-
полного доминирования;
-
неполного доминирования;
-
кодоминирования;
-
сверхдоминирования.
При полном доминировании действие одного гена (доминантного) полностью подавляет действие другого (рецессивного). При скрещивании в первом поколении проявляется доминантный признак (например, желтый цвет горошин).
Рецессивный аллель гена проявляется в результате мутации.
Доминантный аллель отвечает за активную форму фермента, кодирующего признак, а рецессивный аллель - за неактивную форму, или вообще не кодирует белок. У рецессивной особи, гомозиготной по данному аллелю, белок не образуется, и поэтому признак в первом поколении не проявляется.
При неполном доминировании действие рецессивного гена проявляется в первом поколении. Например, при скрещивании растений ночной красавицы с красными и белыми цветами появляются розовые. При этом один ген не обеспечивает достаточное количество белкового продукта для нормального проявления признака.
При кодоминировании проявляется действие обоих генов при одновременном их присутствии. Каждый из аллельных генов кодирует определенный белок. У гетерозиготного организма синтезируются оба белка, и в результате проявляется новый признак. Например, группы крови у человека определяются множественными аллелями IA, IB, I0. Гены IA и IB доминантны, а ген I0 - рецессивен. При взаимодействии генов IA и IB проявляется новый признак, обусловливающий появление IV группы крови IAIB у человека.
При сверхдоминировании у доминантного аллеля в гетерозиготном состоянии отмечается более сильное проявление признака, чем в гомозиготном. У мушки дрозофилы известна рецессивная летальная мутация. Гетерозиготные организмы обладают большей жизнеспособностью, чем доминантные гомозиготные мухи дикого типа.
ВЗАИМОДЕЙСТВИЕ НЕАЛЛЕЛЬНЫХ ГЕНОВ
Очень часто на один признак организма могут оказывать влияние несколько пар неаллельных генов. Это полигенное наследование. Примером полигенного наследования могут служить различные формы взаимодействия неаллельных генов. Взаимодействие неаллельных генов происходит по типу комплементарности,
эпистаза, полимерии.
Комплементарное действие проявляется при одновременном присутствии в генотипе организмов двух доминантных неаллельных генов. Каждый из доминантных генов может проявляться самостоятельно, если другой находится в рецессивном состоянии, но их совместное присутствие в зиготе обусловливает новое состояние признака (рис. 144).
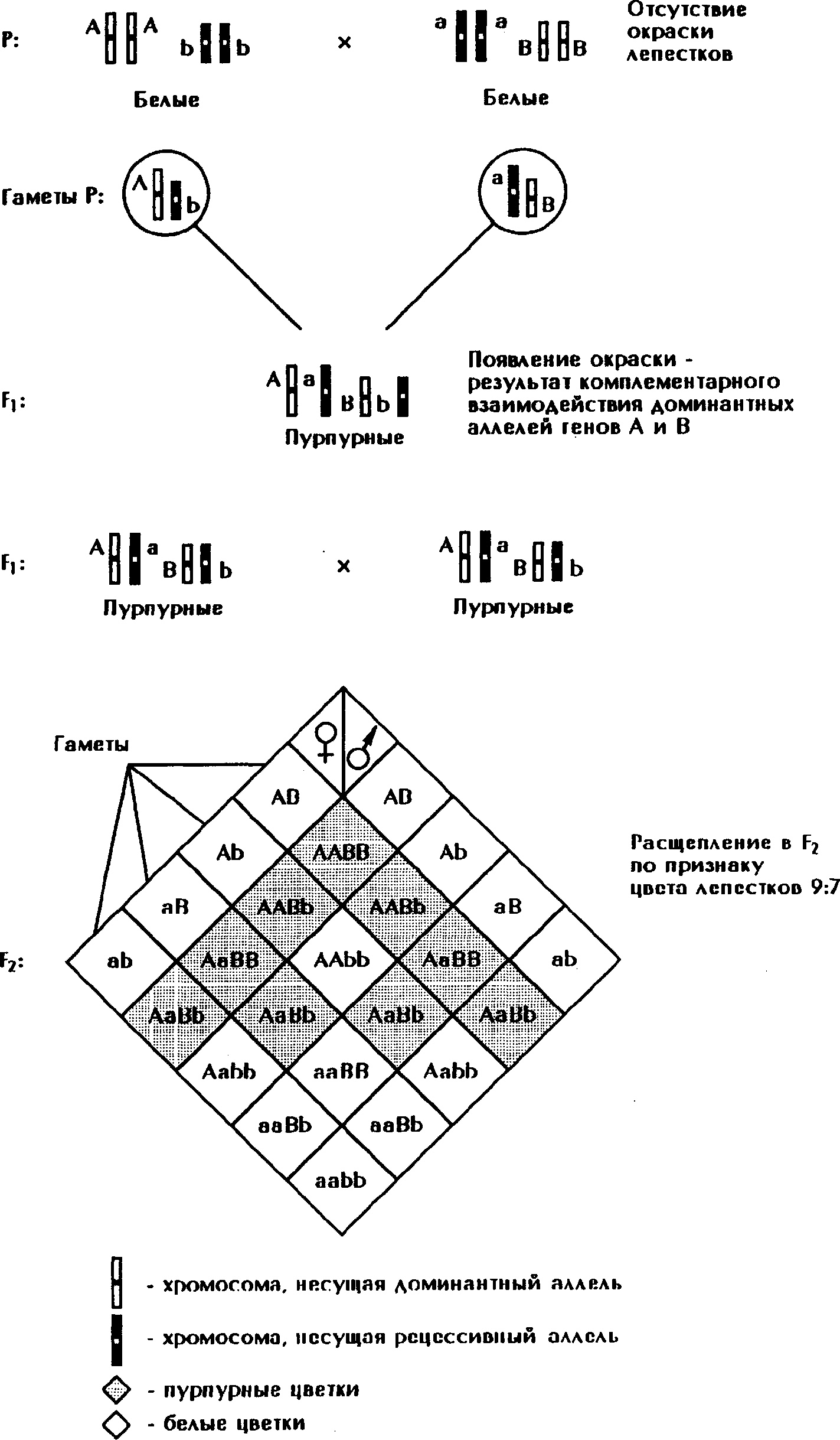
Рис. 144. Комплементарное взаимодействие генов (наследование признака окраски лепестков у душистого горошка). А - аллель, отвечающий за синтез пропигмента (неактивной формы пигмента), а - аллель, не обеспечивающий синтез пропигмента, В - аллель, отвечающий за синтез фермента, который превращает пропигмента в пурпурный пигмента, b - аллель, не обеспечивающий синтез фермента.
Например, скрещивали два сорта душистого горошка с белыми цветами. Гибриды 1-го поколения, полученные в результате скрещивания, имели красные цветы. Окраска цветов зависела от двух взаимодействующих генов.
Белки (ферменты), синтезируемые на основе генов А и В, катализировали биохимические реакции, которые привели к появлению нового признака.
Ген А обусловливал синтез бесцветного предшественника (пропигмента). Ген В определял синтез фермента, под действием которого из пропигмента образовывался пигмент, ответственный за окраску лепестков, а - аллель, не обеспечивающий синтез пропигмента, b - аллель, не обеспечивающий синтез фермента. Лепестки душистого горошка с генотипами ааВВ, ааВв, Аавв, аавв имели белый цвет. Во всех остальных генотипах присутствовали оба доминантных неаллельных гена, что обусловливало образование пропигмента и фермента, участвовавшего в образовании красного пигмента.

При скрещивании гибридов между собой из 16 полученных особей расщепление по генотипу - 9:7. Девять особей имели доминантные гены А и В и были окрашены. Три особи имели доминантный ген А и рецессивный ген в и по фенотипу - белые. Три особи имели рецессивный ген а и доминантный ген В и были белыми. Одна особь, рецессивная по двум генам a и b , имела белую окраску цветов.
Эпистаз - взаимодействие, при котором один из доминантных или рецессивных неаллельных генов подавляет действие другого неаллельного гена. Ген, подавляющий действие другого, называют эпистатическим геном или супрессором. Подавляемый ген называют гипостатическим. Эпистаз бывает доминантным и рецессивным.
Примером доминантного эпистаза является наследование окраски оперения у кур. Доминантный ген С отвечает за развитие окраски оперения у кур. Доминантный неаллельный ген обладает супрессорным действием. В результате этого куры, содержащие в генотипе ген С, в присутствии гена I имеют белое оперение: IICC, IiCC, IiCc, Iicc. Белая окраска оперения обусловлена присутствием рецессивных генов пес или наличием гена - подавителя окраски I. Куры с генотипами iiCC, iiCc будут окрашены (рис. 145).

Рис. 145. Эпистаз. Наследование окраски у кур при взаимодействии двух пар генов. Ген I
подавляет окраску, i - не подавляет ее, С - наличие пигмента, с - отсутствие пигмента.
В основе взаимодействия генов лежат биохимические связи между белками- ферментами, кодируемыми эпистатическими генами.
Эпистатическим действием рецессивного гена можно объяснить бомбейский феномен - необычное наследование антигенов системы групп крови АВ0. Известны четыре группы крови.
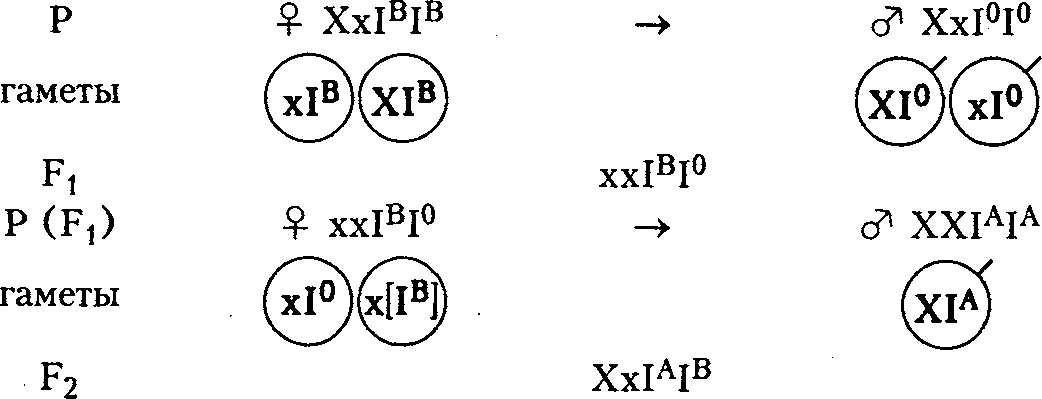 В семье женщины с I группой крови (I0I0) от мужчины со II группой крови (IAIA) родился ребенок с IV группой крови (IAIB), что невозможно и требовало объяснений. При исследовании оказалось, что женщина унаследовала от матери ген IB, а от отца ген I0. Проявлял свое действие только ген I0, и поэтому считалось, что женщина имеет I группу крови. Ген IB был подавлен рецессивным геном х, находящимся в гомозиготном состоянии - хх.
В семье женщины с I группой крови (I0I0) от мужчины со II группой крови (IAIA) родился ребенок с IV группой крови (IAIB), что невозможно и требовало объяснений. При исследовании оказалось, что женщина унаследовала от матери ген IB, а от отца ген I0. Проявлял свое действие только ген I0, и поэтому считалось, что женщина имеет I группу крови. Ген IB был подавлен рецессивным геном х, находящимся в гомозиготном состоянии - хх.
Подавленный ген IB проявил свое действие, и ребенок имел IV группу крови.
Полимерное действие генов связано с тем, что несколько неаллельных генов могут отвечать за один и тот же признак, усиливая его проявление.
Признаки, зависящие от полимерных генов, относят к количественным. Гены, отвечающие за развитие количественных признаков, обладают суммарным эффектом.
Степень проявления признака зависит от числа доминантных аллелей. Чем больше доминантных генов, тем сильнее проявляется признак.
Неаллельные гены отвечают за реализацию одного и того же признака, и поэтому их обозначают одной и той же буквой, цифрами указывая число аллельных пар.
Например, за пигментацию кожи у человека отвечают полимерные неаллельные гены S1 и S2. В присутствии доминантных аллелей этих генов синтезируется много пигмента, рецессивных - мало. Интенсивность пигментации кожи зависит от количества пигмента и определяется количеством доминантных генов.
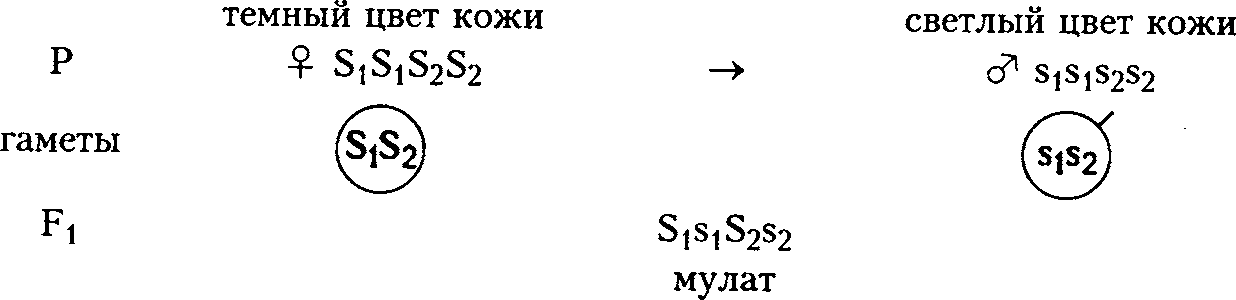 От брака между женщиной с черным цветом кожи и мужчиной с белой кожей родятся мулаты, имеющие промежуточную окраску кожи.
От брака между женщиной с черным цветом кожи и мужчиной с белой кожей родятся мулаты, имеющие промежуточную окраску кожи.
От брака между мулатами с генотипом S1S1S2S2 могут рождаться дети с пигментацией кожи от светлой до темной:
Р ♀ S1S1S2S2 ♂S1S1S2S2
| Гаметы |
|
| ♂ |
| |
| ♀ | S1S2 | S1S2 |
| S1S2 | S1S2 |
| S1S2 S1S2 S1S2 S1S2 | S1S1S2S2 S1S1S2S2 S1S1S2S2 S1S1S2S2 | S1S1S2S2 S1S1S2S2 S1S1S2S2 S1S1S2S2 |
| S1S1S2S2 S1S1S2S2 S1S1S2S2 S1S1S2S2 | S1S1S2S2 S1S1S2S2 S1S1S2S2 S1S1S2S2 |
Вероятность рождения ребенка с белым и черным цветом кожи равна 1/16.
Многие количественные признаки наследуются по принципу полимерии: рост, масса и другие.













Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!
Полезное для учителя

Реализация образовательных программ осуществляется с применением исключительно электронного обучения и ДОТ





