Россия, Каспийск


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Была в сети 03.12.2025 20:16

Арсланалиева Зарема Айсакаевна
учитель экологии и биологии
50 лет
Местоположение
Специализация
Биогеоценозы (экосистемы)
Категория:
Экология
26.10.2025 23:48

Просмотр содержимого документа
«Биогеоценозы (экосистемы)»
66
ТЕМА 5. БИОГЕОЦЕНОЗЫ (ЭКОСИСТЕМЫ)
1. Определение понятия “биогеоценоз” и «экосистема»
2. Структурная и функциональная организация экосистем
2.1. Структурная организация экосистем
2.2. Функциональная организация экосистем
3. Энергетика экосистем и их продуктивность
3.1. Поток энергии и круговорот веществ в экосистемах
3.2. Трофические цепи и сети питания
3.3. Экологическая пирамида
3.4. Биологическая продуктивность экосистем
4. Динамика и стабильность экосистем
4.1. Суточная и годовая динамика экосистем
4.2. Экологические сукцессии
4.3. Стабильные и нестабильные экосистемы
5. Основные типы экосистем
5.1. Наземные экосистемы
5.2. Водные экосистемы
5.3. Различия водных и наземных экосистем
1. Определение понятий «биогеоценоз» и «экосистема»
Биоценоз образует с биотопом биологическую макросистему еще болеет высокого ранга – биогеоценоз.
Термин «биогеоценоз» был предложен в 1940 году русским экологом В.Н. Сукачевым.
Биогеоценоз (от греч. bios – жизнь, geo – земля, koinos – общий) – это однородный участок земной поверхности с определенным составом живых организмов (биоценоз) и условиями среды обитания (биотоп), объединенных обменом веществ и энергии в единый природный комплекс.
В зарубежных странах такие природные комплексы называют экологическими системами (экосистемами).
Термин «экосистема» был предложен в 1935 году английским экологом А. Тенсли.
Экосистема (от греч. oikos – дом, жилище, местообитание, systema - сочетание, объединение) – это безразмерная устойчивая система живых и неживых компонентов, в которой совершается внешний и внутренний круговорот вещества и энергии.
«Экосистема» и «биогеоценоз» - понятия сходные, но не тождественные.
В обоих случаях это устойчивые, взаимодействующие системы живых организмов и среды, в которых совершается поток энергии и круговорот веществ.
Однако, «экосистема» является понятием безразмерным и применяется как к простым системам (капля воды с ее микробным населением, муравейник и др.), так и к сложным искусственным (аквариум, водохранилище, пшеничное поле, космический корабль и др.) и естественным комплексам, какими являются биогеоценозы (тундра, леса, степи, луга, реки, озера, болота и др.).
Биогеоценоз отличается от экосистемы определенностью своего объема, так как всегда связан с определенной частью земной поверхности.
Поэтому, если экосистема может охватывать пространство любой протяженности, - от капли воды с ее микробным населением до биосферы в целом, то биогеоценоз пространственно определяется границами растительного сообщества – фитоценоза.
Иными словами, биогеоценоз – это экосистема в границах фитоценоза.
Следовательно, понятие «экосистема» шире, чем «биогеоценоз», т.е. любой биогеоценоз является экологической системой, но не всякая экосистема может считаться биогеоценозом.
Для обозначения элементарной природной экосистемы используется термин «биогеоценоз».
Природные экосистемы (биогеоценозы) могут формироваться на любом участке земной поверхности.
Экосистемы суши называются наземными экосистемами или биомами.
Биомы – это основные типы наземных экосистем, отличающиеся характером растительности.
По биомной классификации обычно выделяют такие наземные биомы:
тундра (арктическая и альпийская)
хвойные леса
листопадный лес умеренной зоны
степь умеренной зоны
тропическая саванна
пустыни (травянистая и кустарниковая)
вечнозеленый тропический дождевой лес
Экосистемы гидросферы называются водными экосистемами.
В водных экосистемах, где растительность малозаметна, в основе выделения экосистем находятся главные физические характеристики среды (пресные и соленые воды, стоячие и текучие воды).
Водные экосистемы можно классифицировать по следующим признакам:
Пресноводные экосистемы, к которым относятся:
лентические (стоячие воды): озера, пруды, водохранилища и т.д.
лотические (текучие) воды): реки, ручьи и т.д.
заболоченные угодья: болота, болотистые леса.
Морские экосистемы, к которым относятся:
открытый океан (пелагическая зона)
воды континентального шельфа (прибрежные воды)
эстуарии (прибрежные бухты, проливы и устья рек, где смешивается соленая и пресная вода)
морские побережья
Все природные экосистемы (биогеоценозы) связаны между собой и вместе образуют живую оболочку Земли – биосферу, которую можно рассматривать как глобальную экосистему Земли.
Природные экосистемы (биогеоценозы) обычно не имеют четких границ и переходят друг в друга постепенно, образуя довольно обширную переходную (пограничную) зону, которая называется экотоном.
Примерами экотонов для наземных экосистем могут служить:
переходные зоны между лесом и лугом (опушка леса), между лесом и степью (лесостепь), между лесом и тундрой (лесотундра), между лесами с различными видами-эдификаторами (смешанные леса) и др.
Промежуточное положение между наземными и водными экосистемами занимают:
эстуарии (устья рек или заливы океанов, где смешивается соленая и пресная вода), морские побережья, подвергающиеся воздействию приливов и отливов;
прибрежные и внутренние переувлажненные земли (болота, степные блюдца и др.), которые временно или постоянно заполнены водой.
Экотон, занимая промежуточное положение между экосистемами, отличается особыми условиями. Он включает в себя представителей видов растений и животных обеих смежных экосистем, а также нередко такие виды живых организмов, которые не встречаются в этих экосистемах.
В результате экотон обладает большим разнообразием живых организмов, чем близлежащие территории.
В условиях научно-технического прогресса деятельность человека преобразует природные экосистемы (биогеоценозы), что приводит к появлению искусственных экосистем, к которым относятся:
Агроэкосистемы (агробиогеоценозы или агроценозы) – это сообщество живых организмов, созданное для получения сельскохозяйственной продукции и регулярно поддерживаемое человеком.
Агроэкосистемы характеризуются незначительным количеством видов, но высокой их численностью.
Примерами агроэкосистем являются сельскохозяйственные поля, огороды, сады, парки, лесные насаждения, пастбища, пруды и водохранилища, каналы и осушенные болота и др.
Нообиогеоценозы (нооценозы) – это природно-промышленные системы, возникающие в результате промышленной деятельности человека (топливно-энергетические комплексы, промышленные предприятия и др.).
Эколого-экономическая система – это ограниченная определенной территорией часть техносферы, в которой природные, социальные и производственные структуры и процессы связаны взаимоподдерживающими потоками вещества, энергии и информации.
Урбоэкосистемы (урбобиогеоценозы) – это искусственные экосистемы, возникающие в результате поселений человека (крупные города, поселки и др.).
В настоящее время изучению структуры и динамики искусственных экосистем уделяется большое внимание, так как количество их на Земле постоянно увеличивается и уже в обозримом будущем естественных экосистем практически не останется.
2. Структурная и функциональная организация экосистем
2.1. Структурная организация экосистем
В экологии для анализа системы в качестве объекта исследования выбирают элементарную структурную единицу, которая подвергается всестороннему изучению. Необходимым условием построения структурной единицы является то, чтобы она сохраняла все свойства системы.
Понятие “система” означает совокупность не случайно оказавшихся вместе, а составляющих единое целое взаимосвязанных, взаимовлияющих, взаимозависимых компонентов.
Для природных экосистем в качестве объекта исследования принимается биогеоценоз, структурная схема которого представлена на рис.1.
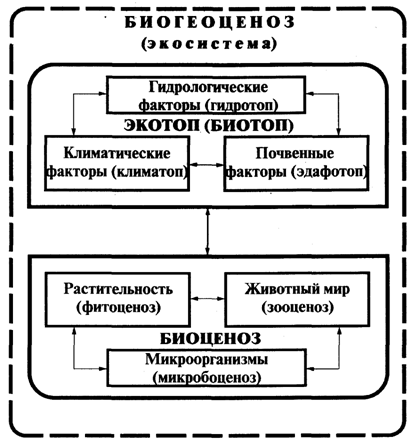
Рис.1. Схема биогеоценоза (экосистемы), по В.Н.Сукачеву
В соответствии со структурной схемой в состав биогеоценоза входят два основных блока:
биотоп - совокупность абиотических факторов среды или весь комплекс факторов неживой природы;
(экотоп – термин, близкий к биотопу, но с подчеркиванием внешних по отношению к сообществу факторов среды, не только абиотических, но и биотических)
биоценоз - совокупность живых организмов.
Биотоп, в свою очередь состоит из совокупности климатических (климатоп) и почвенно-грунтовых (эдафотоп) и гидрологических (гидротоп) факторов среды.
Биоценоз включает сообщества растений (фитоценоз), животных (зооценоз) и микроорганизмов (микробоценоз).
Стрелки на рис.1 обозначают каналы передачи информации между различными компонентами биогеоценоза.
Одним из важнейших свойств биогеоценоза является взаимосвязь и взаимозависимость всех его компонентов.
Вполне понятно, что климат всецело обусловливает состояние и режим почвенно-грунтовых факторов, создает среду обитания для живых организмов.
В свою очередь, почва в какой-то мере определяет климатические особенности (например, от окраски поверхности почвы зависит ее отражательная способность (альбедо), а, следовательно, прогреваемость и влажность воздуха), а также влияет на животных, растения и микроорганизмы.
Все живые организмы теснейшим образом связаны между собой различными пищевыми, пространственными или средообразующими взаимоотношениями, являясь друг для друга либо источником пищи, либо средой обитания, либо фактором смертности.
Особенно важна роль микроорганизмов (в первую очередь бактерий) в процессах почвообразования, минерализации органических веществ и нередко выступающих в качестве возбудителей заболеваний растений и животных.
2.2. Функциональная организация экосистем.
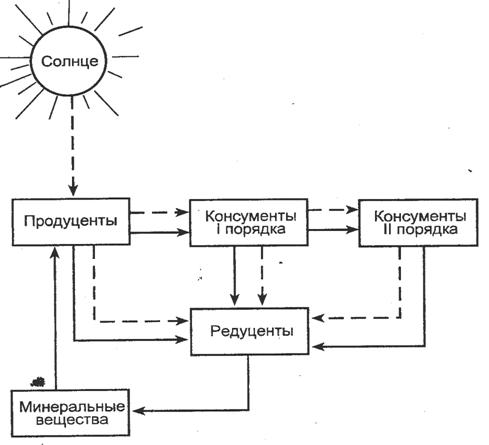
Основная функция экосистем - это поддержание круговорота веществ в биосфере, которая базируется на пищевых взаимоотношениях видов.
Несмотря на огромное многообразие видов, входящих в состав различных сообществ, каждая экосистема с необходимостью включает представителей трех функциональных групп организмов - продуцентов, консументов и редуцентов.
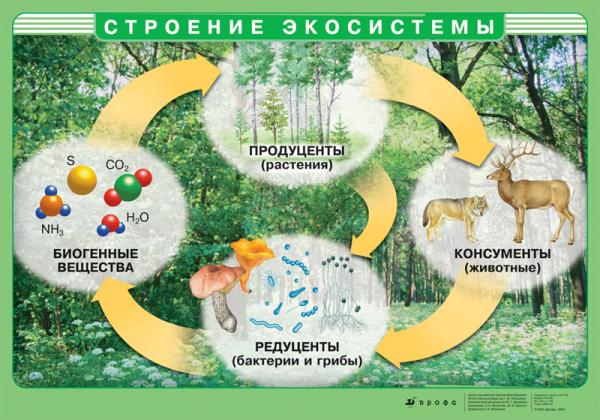
Основу подавляющего большинства биогеоценозов составляют продуценты (производители) - это автотрофные организмы (от греч. “авто” - сам и “трофо” - питание), которые обладают способностью синтезировать органические вещества из неорганических, используя для этого солнечную энергию или энергию химических связей.
В зависимости от источника используемой энергии различают два типа организмов: фотоавтотрофы и хемоавтотрофы.
Фотоавтотрофы - это организмы, которые с использованием солнечной энергии способны создавать органические вещества в процессе фотосинтеза.
К фотоавтотрофным организмам относятся растения, а также сине-зеленые водоросли (цианобактерии).
Однако не все растения являются продуцентами, например:
некоторые грибы (шляпочные, плесневые), а также некоторые цветковые виды (например, подъельник), которые не содержат хлорофилла, не способны к фотосинтезу и поэтому питаются готовыми органическими веществами.
Хемоавтотрофы - это организмы, которые в качестве источника энергии для образования органических веществ используют энергию химических связей.
К хемоавтотрофным организмам относятся: водородные, нитрифицирующие бактерии, железобактерии и др.
Группа организмов-хемоавтотрофов немногочисленна и не играет принципиальной роли в биосфере.
Только продуценты (производители) способны сами производить для себя богатую энергией пищу, т.е. являются самостоятельно питающимися. Более того, они непосредственно или косвенно обеспечивают питательными элементами консументы и редуценты.
Консументы (потребители) - это гетеротрофные организмы (от греч. “гетеро” - разный), которые используют живое органическое вещество в качестве пищи для получения и накопления энергии.
Основным источником энергии для гетеротрофных организмов является энергия, выделяющаяся при расщеплении химических связей органических веществ, созданных автотрофными организмами.
Таким образом, гетеротрофы всецело зависят от автотрофов.
В зависимости от источников питания различают:
Консументы первого порядка (фитофаги) - это растительноядные организмы, питающиеся разными видами растительного корма (продуцентами).
Примерами первичных консументов являются:
птицы едят семена, почки и листву;
олени и зайцы питаются ветками и листьями;
кузнечики и многие другие виды насекомых потребляют в пищу все части растений;
в водных экосистемах зоопланктон (мелкие животные, передвигающиеся преимущественно с течением воды) питается фитопланктоном (микроскопические, обычно одноклеточные водоросли).
Консументы второго порядка (зоофаги) - это плотоядные организмы, питающиеся исключительно растительноядными организмами (фитофагами).
Примерами вторичных консументов являются:
насекомоядные млекопитающие, птицы и пауки, поедающие насекомых;
чайки, поедающие моллюсков и крабов;
лиса, питающаяся зайцами;
тунец, питающийся сельдью и анчоусами.
Консументы третьего порядка - это хищники, питающиеся только плотоядными организмами.
Примерами третичных консументов являются:
ястреб или сокол, питающийся змеями и горностаями;
акулы, питающиеся другими рыбами.
Встречаются консументы четвертого и более высоких порядков.
Кроме того, существует много видов со смешанным типом питания:
когда человек ест фрукты и овощи, то он является консументом первого порядка;
когда человек питается мясом растительноядного животного, то он вторичный консумент;
когда человек ест рыбу, питающуюся другими животными, которые в свою очередь употребляют в пищу водоросли, то человек выступает в роли консумента третьего порядка.
Эврифаги - это всеядные организмы, которые питаются как растительной, так и животной пищей.
Например: свиньи, крысы, лисы, тараканы и человек.
Редуценты (разрушители) - это гетеротрофные организмы, которые питаются мертвым органическим веществом и минерализуют его до простых неорганических соединений.
Различают два основных типа редуцентов: детритофаги и деструкторы.
Детритофаги - это организмы, которые непосредственно потребляют мертвые растительные и животные остатки (детрит).
К детритофагам относятся: шакалы, грифы, крабы, термиты, муравьи, дождевые черви, многоножки и др.
Деструкторы - это организмы, которые разлагают сложные органические соединения мертвой материи до более простых неорганических веществ, которые затем используются продуцентами.
Основными деструкторами являются: бактерии и грибы.
При этом бактерии принимают участие в разложении остатков животного происхождения, так как тяготеют к субстратам со слабощелочной реакцией.
А грибы, наоборот, предпочитают слабокислые субстраты, поэтому они принимают основное участие в разложении остатков растительного происхождения.
Таким образом, каждый живой организм в составе биогеоценоза выполняет определенную функцию, т.е. занимает определенную экологическую нишу в сложной системе экологических взаимоотношений с другими организмами и факторами неживой природы.
Так, например, в разных частях света и на разных территориях встречаются неодинаковые в систематическом отношении, но экологически сходные виды которые выполняют одинаковые функции в своих биогеоценозах:
травянистая и лесная растительность Австралии по видовому составу существенно отличается от растительности сходного климатического района Европы или Азии, но как продуценты в своих биогеоценозах они выполняют одинаковые функции, т.е. занимают в принципе одинаковые экологические ниши;
антилопы в саваннах Африки, бизоны в прериях Америки, кенгуру в саваннах Австралии, являясь консументами первого порядка, выполняют одинаковые функции, т.е. занимают сходные экологические ниши в своих биогеоценозах.
Вместе с тем, часто близкие в систематическом отношении виды, поселяясь рядом в одном биогеоценозе, выполняют неодинаковые функции, т.е. занимают разные экологические ниши:
два вида водяных клопов в одном и том же водоеме играют различную роль: один вид ведет хищнический образ жизни и является третичным консументом, а другой питается мертвыми и разлагающимися организмами и является редуцентом. Это приводит к уменьшению конкурентного напряжения между ними.
Кроме того, один и тот же вид в разные периоды своего развития может выполнять различные функции, т.е. занимать различные экологические ниши:
головастик питается растительной пищей и является первичным консументом, а взрослая лягушка - типичное плотоядное животное, является консументом второго порядка;
среди водорослей имеются виды, которые функционируют то как автотрофы, то как гетеротрофы. В результате в определенные периоды своей жизни они выполняют различные функции и занимают те или иные экологические ниши.
3. Энергетика экосистем и их продуктивность.
3.1. Поток энергии и круговорот веществ в экосистемах.
Все живые организмы в процессе своей жизнедеятельности потребляют и рассеивают энергию. Поэтому ни один живой организм не может существовать в течение сколько-нибудь продолжительного времени без постоянного притока свободной энергии извне.
Главным источником такой свободной энергии для подавляющего большинства живых организмов на Земле является энергия солнечного света.
Фотосинтезирующие организмы непосредственно используют энергию солнечного света. При этом в процессе фотосинтеза из диоксида углерода и воды образуются сложные органические вещества, в которых часть солнечной энергии запасена в форме химических связей, и в качестве побочного продукта выделяется кислород:
6 СО2 + 6 Н2О + солнечная энергия = С6Н12О6 + 6 О2
Полученные органические вещества могут служить источником энергии не только для самого растения, но и для других организмов в биогеоценозе.
Высвобождение заключенной в органических веществах энергии происходит в процессе клеточного дыхания.
Клеточное дыхание - это процесс постепенного сгорания (окисления) органических соединений, находящихся в клетках живых организмов:
С6Н12О6 + 6 О2 = 6 СО2 + 6 Н2О + энергия
Продукты клеточного дыхания - диоксид углерода и вода могут вновь использоваться растениями для получения органических веществ в процессе фотосинтеза.
Таким образом, в экосистеме совершается бесконечный круговорот веществ (рис.2).
Однако, содержащаяся в органических веществах энергия не совершает круговорот, а постепенно превращается в бесполезную тепловую энергию, которая рассеивается в пространстве, так как согласно второму закону термодинамики переход энергии из одной формы (например, энергии света) в другую (например, потенциальную энергию пищи) сопровождается снижением количества полезной энергии.
Поэтому необходимым условием существования экосистемы является постоянный приток энергии извне.


Процесс фотосинтеза
6 СО2 + 6 Н2О + солнечная энергия = С6Н12О6 + 6 О2




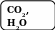



Процесс клеточного дыхания
C6H12O6 + 6 О2 = 6 СО2 + 6 Н2О + энергия

Рис. 2. Суммарный поток энергии и круговорот веществ в экосистемах
поток энергии – светлые стрелки
круговорот веществ – темные стрелки
Для биологического круговорота веществ необходимо наличие в экосистемах:
органимов-продуцентов, которые используют энергию излучения Солнца для превращения неорганических веществ в более сложные органические соединения, в которых часть солнечной энергии запасена в форме химических связей;
организмов-консументов, которые используют эти органические вещества для получения и накопления энергии;
организмов-редуцентов (деструкторов), которые в процессе своей жизнедеятельности разлагают органические соединения на более простые неорганические вещества.
На рис.3 показана связь основных структурных компонентов экосистемы между собой потоком энергии и круговоротом веществ.
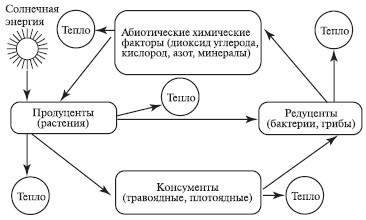
Рис.3. Связь основных структурных компонентов экосистемы между собой
потоком энергии и круговоротом веществ.
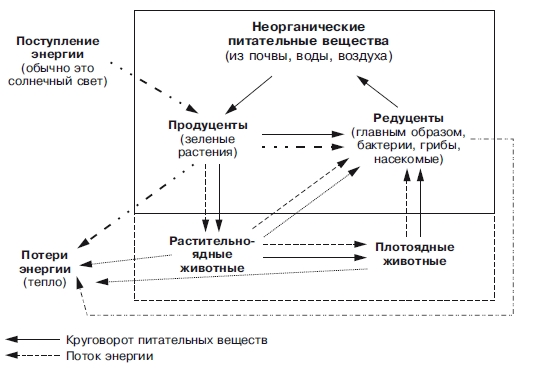
Продуценты поглощают около 1% падающей на Землю солнечной энергии и используют ее для синтеза органических соединений, в которых солнечная энергия запасена в форме химических связей.
Более половины этой энергии расходуется продуцентами на различные процессы жизнедеятельности, превращаясь при этом в теплоту и рассеиваясь в пространстве.
Остальная часть энергии запасается в виде вновь возникшей биомассы растений и может быть частично использована гетеротрофными организмами-консументами в качестве пищи.
Когда растительноядное животное поедает растения, то большая часть энергии, которая содержится в пище, используется на различные процессы жизнедеятельности, превращаясь при этом в теплоту и рассеиваясь в пространстве.
Только 5 - 20% энергии пищи переходит во вновь созданную биомассу растительноядных животных.
Если растительноядное животное съедается плотоядным хищником, то вновь теряется большая часть заключенной в пище энергии, которая расходуется на различные процессы жизнедеятельности плотоядного животного, а лишь немногая часть переходит в их биомассу.
Часть биомассы продуцентов, не съеденная консументами, отмирает и энергия, аккумулированная в отмершей биомассе, поступает в почву в виде растительного опада.
Растительный и животный опад, а также продукты жизнедеятельности живых организмов являются пищей редуцентов.
При этом большая часть энергии, содержащаяся в пище, рассеивается в виде тепла в результате жизнедеятельности редуцентов, а некоторая часть энергии запасается в биомассе редуцентов.
Редуценты-деструкторы являются завершающим звеном в круговороте веществ, так как сложные органические соединения в мертвых растительных и животных организмах, а также в продуктах жизнедеятельности живых организмов разлагаются редуцентами на более простые неорганические вещества, которые вновь используются продуцентами.
Без редуцентов весь мир вскоре был бы погружен в слой растительного и животного опада и различного мусора.
Приведенная схема свидетельствует о том, что экосистема может существовать без консументов, так как химические вещества могут переходить прямо от продуцентов к редуцентам и обратно без участия консументов.
Кроме того, существуют биогеоценозы, которые не включают автотрофных фотосинтезирующих растений (например, экосистемы больших глубин океана или пещер).
Но во всех экосистемах подобного типа обязателен приток энергии извне в виде органических веществ, в которых аккумулирована солнечная энергия.
Таким образом, если движение вещества, вызванное деятельностью организмов, совершает круговорот, т.е. вещество может быть использовано многократно, то поток энергии имеет однонаправленный характер, т.е. содержащаяся в пище энергия не совершает круговорота, а постепенно превращается в тепловую энергию и рассеивается в пространстве.
Поэтому необходимым условием существования экосистемы является постоянный приток энергии извне, которая необходима всем организмам для поддержания их жизнедеятельности и самовоспроизведения.
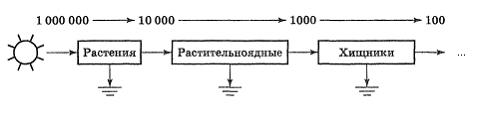
3.2. Трофические цепи и сети питания.
В экосистемах продуценты, консументы и редуценты объединены сложными процессами переноса веществ и энергии, которая заключена в пище, созданной преимущественно растениями.
Перенос потенциальной энергии пищи, созданной растениями, через ряд организмов путем поедания одних видов другими называется трофической (пищевой) цепью, а каждое ее звено называется трофическим уровнем.
Все организмы, пользующиеся одним типом пищи, принадлежат к одному трофическому уровню.
На рис.4. представлена схема трофической цепи.
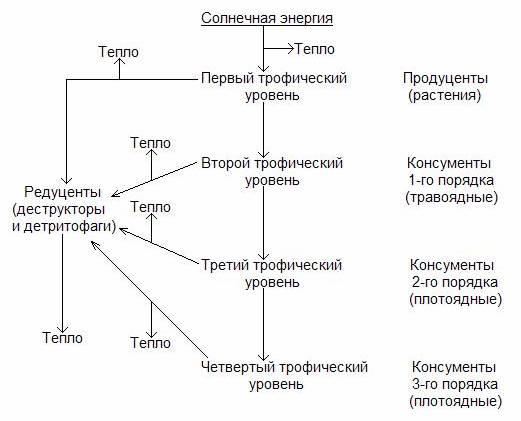
Рис.4. Схема пищевой цепи.
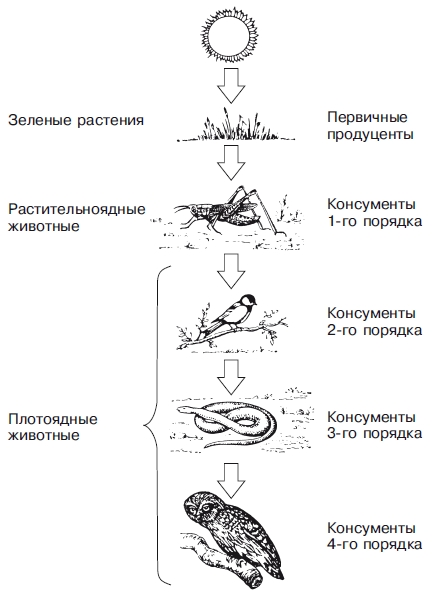
Рис.4. Схема пищевой цепи.
Первый трофический уровень образуют продуценты (зеленые растения), которые аккумулируют солнечную энергию и создают органические вещества в процессе фотосинтеза.
При этом более половины энергии, запасенной в органических веществах, расходуется в процессах жизнедеятельности растений, превращаясь при этом в тепло и рассеиваясь в пространстве, а остальная часть поступает в пищевые цепи и может быть использована гетеротрофными организмами последующих трофических уровней при питании.
Второй трофический уровень образуют консументы 1-го порядка - это растительноядные организмы (фитофаги), которые питаются продуцентами.
Консументы первого порядка большую часть энергии, которая содержится в пище, расходуют на обеспечение своих жизненных процессов, а остальную часть энергии используют на построение собственного тела, преобразуя тем самым растительные ткани в животные.
Таким образом, консументы 1-го порядка осуществляют первый, принципиальный этап трансформации органического вещества, синтезированного продуцентами.
Первичные консументы могут служить источником питания для консументов 2-го порядка.
Третий трофический уровень образуют консументы 2-го порядка - это плотоядные организмы (зоофаги), которые питаются исключительно растительноядными организмами (фитофагами).
Консументы 2-го порядка осуществляют второй этап трансформации органического вещества в цепях питания.
Однако, химические вещества, из которых строятся ткани животных организмов, довольно однородны и поэтому трансформация органического вещества при переходе со второго трофического уровня консументов на третий не имеет столь принципиального характера, как при переходе с первого трофического уровня на второй, где происходит преобразование растительных тканей в животные.
Вторичные консументы могут служить источником питания для консументов 3-го порядка.
Четвертый трофический уровень образуют консументы 3-го порядка - это плотоядные животные, питающиеся только плотоядными организмами.
Последний уровень трофической цепи занимают редуценты (деструкторы и детритофаги).
Редуценты-деструкторы (бактерии, грибы, простейшие) в процессе своей жизнедеятельности разлагают органические остатки всех трофических уровней продуцентов и консументов до минеральных веществ, которые вновь возвращаются к продуцентам.
Все звенья трофической цепи взаимосвязаны и взаимозависимы.
Между ними от первого к последнему звену осуществляется передача веществ и энергии. Однако, необходимо отметить, что при передаче энергии с одного трофического уровня на другой происходит ее потеря. В результате цепь питания не может быть длинной и чаще всего состоит из 4-6 звеньев.
Однако, такие пищевые цепи в чистом виде в природе обычно не встречаются, поскольку каждый организм имеет несколько источников питания, т.е. пользуется несколькими типами пищи, и сам используется как продукт питания другими многочисленными организмами из одной и той же трофической цепи или даже из разных цепей питания.
Например:
паразиты могут использовать в качестве источника питания различные виды растений и животных из разных трофических уровней;
всеядные организмы потребляют в пищу как продуцентов, так и консументов, т.е. являются одновременно консументами первого, второго, а иногда и третьего порядка;
комар, питающийся кровью человека и хищных животных, находится на очень высоком трофическом уровне. Но комарами питается болотное растение росянка, которая, таким образом, является и продуцентом и консументом высокого порядка.
Поэтому, практически любой организм, входящий в состав одной трофической цепи, одновременно может входить и в состав других трофических цепей.
Таким образом, трофические цепи могут многократно разветвляться и переплетаться, образуя сложные сети питания или трофические (пищевые) сети, в которых многочисленность и разнообразие пищевых связей выступает как важный механизм поддержания целостности и функциональной устойчивости экосистем.
На рис.5. показана упрощенная схема сети питания для наземной экосистемы.
Вмешательство человека в природные сообщества организмов путем намеренного или ненамеренного устранения какого-либо вида часто имеет непредсказуемые негативные последствия и приводит к нарушению устойчивости экосистем.
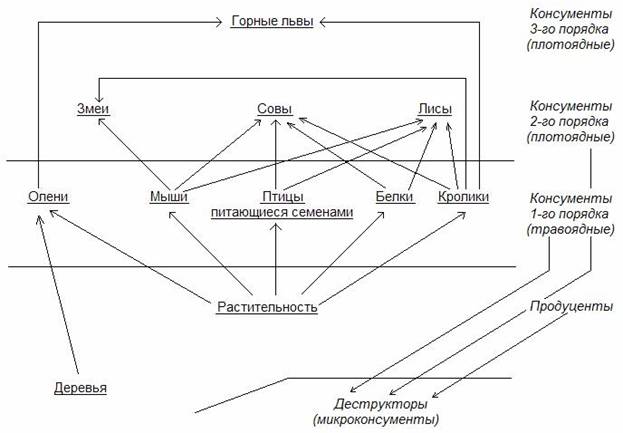
Рис.5. Схема трофической сети.
Рис.5. Схема трофической сети.
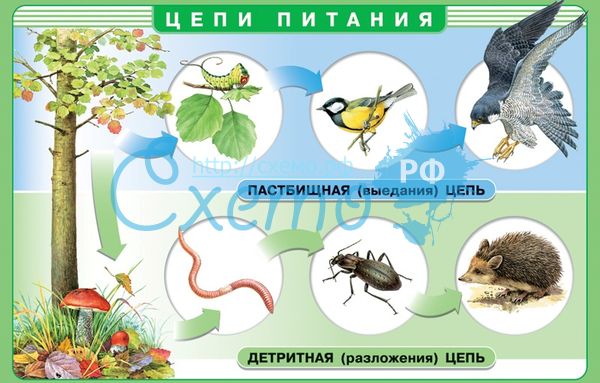
Существует два основных типа трофических цепей:
пастбищные цепи (цепи выедания или или цепи потребления);
детритные цепи (цепи разложения).
Пастбищные цепи (цепи выедания или цепи потребления) - это процессы синтеза и трансформации органических веществ в трофических цепях.
Пастбищные цепи начинаются с продуцентов. Живые растения поедаются фитофагами (консументами первого порядка), а сами фитофаги являются пищей для плотоядных животных (консументов второго порядка), которыми могут питаться консументы третьего порядка и т.д.
Примеры пастбищных цепей для наземных экосистем:
3 звена: осина → заяц → лиса; растение → овца → человек.
4 звена: растения → кузнечики → ящерицы → ястреб;
нектар цветка растения → муха → насекомоядная птица →
хищная птица.
5 звеньев: растения → кузнечики → лягушки → змеи → орел.
Примеры пастбищных цепей для водных экосистем:→
3 звена: фитопланктон → зоопланктон → рыбы;
5 звеньев: фитопланктон → зоопланктон → рыбы → хищные рыбы →
хищные птицы.
Детритные цепи (цепи разложения) - это процессы поэтапной деструкции и минерализации органических веществ в трофических цепях.
Детритные цепи начинаются с поэтапного разрушения мертвого органического вещества детритофагами, которые последовательно сменяют друг друга в соответствии со специфичным типом питания.
На последних стадиях деструкционных процессов функционируют редуценты-деструкторы, минерализующие остатки органических соединений до простых неорганических веществ, которые вновь используются продуцентами.
Например, при разложении мертвой древесины последовательно сменяют друг друга: жуки → дятлы → муравьи и термиты → грибы-деструкторы.
Детритные цепи наиболее распространены в лесах, где большая часть (около 90%) ежегодного прироста биомассы растений не потребляется непосредственно растительноядными животными, а отмирает и попадает в эти цепи в виде листового опада, подвергаясь затем разложению и минерализации.
В водных экосистемах большая часть вещества и энергии включается в пастбищные цепи, а в наземных экосистемах наибольшее значение имеют детритные цепи.
Таким образом, на уровне консументов происходит разделение потока органического вещества по разным группам потребителей:
живое органическое вещество следует по пастбищным цепям;
мертвое органическое вещество идет по детритным цепям.
3.3. Экологическая пирамида.
В любой трофической цепи не вся пища используется на рост особи, т.е. на накопление ее биомассы. Часть ее расходуется на удовлетворение энергетических затрат организма (дыхание, движение, размножение, поддержание температуры тела).
При этом биомасса одного звена не может быть полностью переработана последующим, и в каждом последующем звене трофической цепи происходит уменьшение биомассы.
В среднем считается, что лишь порядка 10% биомассы и связанной в ней энергии переходит с каждого трофического уровня на следующий, т.е. продукция организмов каждого последующего трофического уровня всегда меньше в среднем в 10 раз продукции предыдущего уровня.
Так, например, в среднем из 1000 кг растений образуется 100 кг биомассы растительноядных животных (консументов первого порядка). Плотоядные животные (консументы второго порядка), поедающие растительноядных, могут синтезировать из этого количества 10 кг своей биомассы, а хищники (консументы третьего порядка), которые питаются плотоядными животными, синтезируют только 1 кг своей биомассы.
Таким образом, суммарная биомасса, заключенная в ней энергия, а также численность особей прогрессивно уменьшаются по мере восхождения по трофическим уровням.
Эта закономерность получила название правила экологической пирамиды.
Данное явление впервые было изучено Ч.Элтоном (1927 г.) и названо им пирамидой чисел или пирамидой Элтона.
Экологическая пирамида - это графическое изображение соотношения между продуцентами и консументами разных порядков, выраженное в единицах биомассы (пирамида биомасс), числа особей (пирамида численности) или заключенной в массе живого вещества энергии (пирамида энергии) (рис.6).
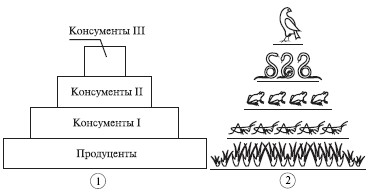
Рис.6. Схема экологической пирамиды.
Экологическая пирамида выражает трофическую структуру экосистем в геометрической форме.
Различают три основных типа экологических пирамид: пирамида чисел (численности), пирамида биомассы и пирамиды энергии.
1) пирамиды чисел, основанные на подсчете организмов каждого трофического уровня;
2) пирамиды биомассы, в которых используется суммарная масса (обычно сухая) организмов на каждом трофическом уровне;
3) пирамиды энергии, учитывающие энергоемкость организмов каждого трофического уровня.
Пирамиды энергии считаются самыми важными, поскольку они непосредственно обращаются к основе пищевых отношений — потоку энергии, необходимой для жизнедеятельности любых организмов.
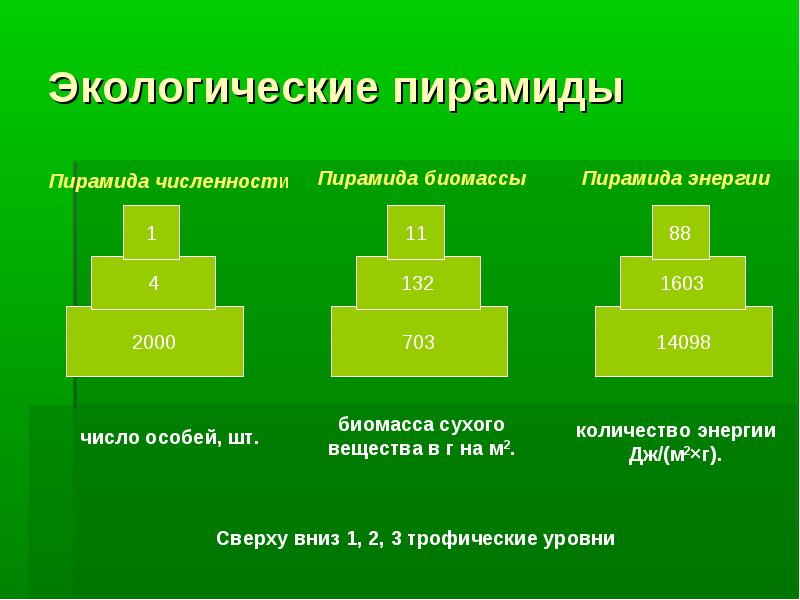
Пирамида чисел (численности)
Пирамида чисел (численности) или пирамида Элтона отражает численность отдельных организмов на каждом трофическом уровне.
Пирамида численности представляет собой наиболее простое приближение к изучению трофической структуры экосистемы.
При этом сначала подсчитывают число организмов на данной территории, сгруппировав их по трофическим уровням и представив в виде прямоугольника, длина (или площадь) которого пропорциональна числу организмов, обитающих на данной площади (или в данном объеме, если это водная экосистема).
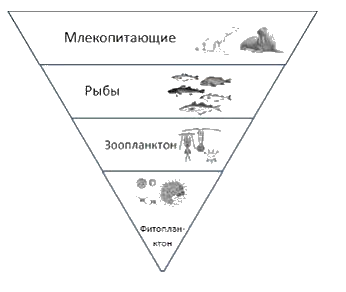
Пирамида численности может иметь правильную форму, т.е. суживаться кверху (правильная или прямая), а может быть и перевернутой вершиной вниз (перевернутая или обращенная) рис.7.

правильная (прямая) перевернутая (обращенная)
(пруд, озеро, луг, степь, пастбище и др.) (лес умеренного пояса летом и др.)
Рис.7. Пирамида численности (1 – правильная; 2- перевернутая)
Пирамида численности имеет правильную форму, т.е. сужается при продвижении от уровня продуцентов к более высоким трофическим уровням, для водных экосистем (пруд, озеро и др.) и наземных экосистем (луг, степь, пастбище и др.).
Например:
тысяча особей фитопланктона в небольшом пруду может прокормить 100 особей мелких ракообразных – консументов первого порядка, которые в свою очередь прокормят 10 особей рыб – консументов второго порядка, которых будет достаточно, чтобы прокормиться 1 окуню – консументу третьего порядка.
Пирамида численности для некоторых экосистем, например для леса умеренного пояса, имеет перевернутую форму.
Например:
в лесу умеренного пояса летом небольшое количество больших деревьев - продуцентов снабжает пищей огромное количество небольших по размеру насекомых-фитофагов и птиц - консументов первого порядка.
Однако в экологии пирамида численности употребляется редко, так как из-за большого числа особей на каждом трофическом уровне очень трудно отобразить структуру биоценоза в одном масштабе.
Пирамида биомассы
Пирамида биомассы отражает более полно пищевые взаимоотношения в экосистеме, так как в ней учитывается суммарная масса организмов (биомасса) каждого трофического уровня.
Прямоугольники в пирамидах биомассы отображают массу организмов каждого трофического уровня, отнесенную к единице площади или объема.
Пирамиды биомассы, так же, как и пирамиды численности, могут быть не только правильной формы, но и перевернутыми (обращенными) рис.8.
 Консументы 3 порядка
Консументы 3 порядка

Консументы 2 порядка

 Консументы 1 порядка
Консументы 1 порядка
П
ро
Продуценты
правильная (прямая) перевернутая (обращенная)
(наземные экосистемы: (водные экосистемы: озеро,
луг, поле и др.) пруд и особенно морские
экосистемы)
Рис.7. Пирамида биомасс (1 – правильная; 2- перевернутая)
Для большинства наземных экосистем (луг, поле и др.) суммарная биомасса каждого последующего трофического уровня пищевой цепи уменьшается.
Это создает пирамиду биомасс, где существенно преобладают продуценты, а над ними располагаются постепенно уменьшающиеся трофические уровни консументов, т.е. пирамида биомасс имеет правильную форму.
Например:
в среднем из 1000 кг растений образуется 100 кг тела растительноядных животных – консументов первого порядка (фитофагов). Плотоядные животные – консументы второго порядка, поедающие растительноядных, могут синтезировать из этого количества 10 кг своей биомассы. А хищники – консументы третьего порядка, питающиеся плотоядными животными, синтезируют только 1 кг своей биомассы.
В водных экосистемах (озеро, пруд и др.) пирамида биомасс может быть перевернутой, где биомасса консументов преобладает над биомассой продуцентов.
Это объясняется тем, что в водных экосистемах продуцентом является микроскопический фитопланктон, быстро растущий и размножающийся), который в достаточном количестве непрерывно поставляет живую пищу консументам, намного медленно растущим и размножающимся. Зоопланктон (или другие животные, питающиеся фитопланктоном) накапливают биомассу годами и десятилетиями, тогда как фитопланктон имеет крайне короткий период жизни (несколько дней или часов).
Пирамида энергии
Пирамида энергии дает наиболее полное представление о функциональной организации сообщества, так как она отражает величину потока энергии, скорость прохождения биомассы через пищевую цепь.
В противоположность пирамидам чисел и биомассы, отражающим статистику экосистемы, т.е. характеризующим количество организмов или их биомассу в данный момент времени, пирамида энергии отражает на каждом уровне удельное количество энергии (на единицу площади или объема), прошедшей через предыдущий трофический уровень за данный отрезок времени, т.е. динамику экосистем.
Р.Линдеман (1942 г.) сформулировал правило пирамиды энергии (правило 10%), согласно которому с одного трофического уровня экологической пирамиды переходит на другой, более высокий, в среднем не более 10% энергии.
Остальная часть энергии теряется в виде теплового излучения в соответствии со вторым законом термодинамики.
Поэтому организмы в результате процессов обмена теряют в каждом звене пищевой цепи около 90% всей энергии, которая расходуется на поддержание их жизнедеятельности.
По этой причине пирамида энергии всегда суживается кверху, т.е. имеет правильную форму рис.9.




10 Консументы 3 порядка 9

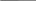
100 Консументы 2 порядка 90


1000 Консументы 1 порядка 900


10000 Продуценты 9000

Полезная энергия, доступная на Энергия, теряемая в виде тепла
каждом трофическом уровне на каждом трофическом уровне
(в ккал) (в ккал)
Рис.9. Пирамиды энергетических потоков и расходов энергии
Пирамиды энергетических потоков и расхода энергии свидетельствуют о том, что чем длиннее пищевая цепь, тем больше теряется полезной энергии.
Поэтому цепи питания не могут быть очень длинными, обычно они состоят из 3-5 звеньев (трофических уровней).
Пирамида энергетических потоков объясняет, почему можно прокормить большее число людей, если сократить пищевую цепь до прямого потребления зерновых («рис-человек»), чем если в качестве пищи использовать животных, потребляющих зерно («зерно-говядина-человек»).
Чтобы избежать белкового (протеинового) голодания, вегетарианское питание должно состоять из разнообразных растений, имеющих в своем составе молекулы 10 азотсодержащих аминокислот, используемых для создания протеина, который наш организм не в состоянии сам синтезировать.
Бедная часть населения нашей планеты, довольствующая растительной диетой, часто не имеет средств для выращивания или покупки всех овощей и фруктов, необходимых для избегания протеинового голодания.
Правило экологической пирамиды универсально и объективно отражает круговорот веществ и поток энергии в биосфере. В масштабе всей биосферы это правило никогда не нарушается.
3.4. Биологическая продуктивность экосистем.
Чтобы оценить значение того или иного вида для круговорота веществ в данном биогеоценозе необходимо знать не только его биомассу, но и относительную скорость ее создания, т.е. биологическую продуктивность.
Таким образом,
Биологическая продуктивность - это скорость создания определенного количества биомассы растений, животных и микроорганизмов, входящих в состав биогеоценоза.
Биологическая продуктивность определяется количеством биомассы, синтезируемой за единицу времени на единицу площади (или объема) и выражается чаще всего в граммах углерода или сухого органического вещества или в энергетических единицах – эквивалентном числе калорий или джоулей.
Биологическую продуктивность можно выразить продукцией за сезон, за год, за несколько лет или за любую другую единицу времени.
Для наземных и донных организмов биологическая продуктивность определяется количеством биомассы на единицу площади, а для планктонных и почвенных - на единицу объема.
Ключевое слово в понятии продуктивность – скорость. Однако вместо термина «продуктивность» часто используется термин «продукция», но при этом все равно учитывается фактор времени.
Биологическую продуктивность нельзя смешивать с биомассой.
Биомасса - это выраженное в единицах массы (веса) или энергии количество живого вещества тех или иных организмов, обитающих на исследуемой площади или в исследуемом объеме.
Например:
планктонные водоросли за год на единицу площади синтезируют столько же органического вещества, сколько и высокопродуктивные леса, однако биомасса последних в сотни тысяч раз больше;
популяции мелких млекопитающих по сравнению с крупными обладают большей скоростью роста и размножения и поэтому имеют более высокую продуктивность при равной биомассе.
Различают первичную и вторичную продуктивность экосистем.
Первичная продуктивность экосистем - это скорость, с которой автотрофные организмы (продуценты) в процессе фотосинтеза связывают солнечную энергию и запасают ее в форме химических связей органических веществ, т.е. скорость образования биомассы органического вещества автотрофами (продуцентами).
Первичная продуктивность подразделяется на валовую и чистую продуктивность.
Валовая первичная продуктивность – это скорость накопления органического вещества продуцентами, включая затраты на дыхание (т.е. включая ту его часть, которая будет израсходована в процессах жизнедеятельности растений).
Так, например, в тропических лесах и зрелых лесах умеренной зоны затраты на дыхание составляют 40-70%, а у планктонных водорослей и у большинства сельскохозяйственных культур – 40%.
Чистая первичная продуктивность – это скорость накопления органического вещества в растительных тканях за вычетом той его части, которая использовалась на дыхание растений.
Поэтому чистая первичная продукция, накопленная в виде биомассы растений всегда меньше валовой первичной продукции, созданной ими в процессе фотосинтеза.
Чистая первичная продуктивность автотрофных организмов (продуцентов) может служить источником питания для гетеротрофных организмов, которые на ее основе образуют свою биомассу.
Вторичная продуктивность - это скорость образования биомассы гетеротрофными организмами (консументами).
Вторичная продуктивность уже не делится на валовую и чистую продуктивность, так как гетеротрофы увеличивают свою массу за счет первичной ранее созданной продукции.
Вторичную продуктивность рассчитывают отдельно для каждого трофического уровня, так как прирост биомассы на каждом из них происходит за счет энергии, поступающей с предыдущего уровня.
При этом необходимо учитывать, что при переходе с одного трофического уровня консументов на другой значительная часть энергии расходуется в процессах жизнедеятельности, поэтому вторичная продукция каждого последующего трофического уровня будет меньше продукции предыдущего.
Если в экосистеме скорость образования чистой первичной продукции выше темпов переработки ее консументами, то это ведет к увеличению биомассы продуцентов.
Если при этом присутствует недостаточная утилизация продуктов опада в цепях разложения редуцентами, то происходит накопление мертвого органического вещества (в форме каменного угля, горючих сланцев, сухих листьев и т.д.).
В стабильных экосистемах биомасса остается постоянной, так как практически вся созданная продукция расходуется в цепях питания разнообразными консументами и редуцентами, т.е. природа стремится использовать полностью валовую продукцию.
Однако равенство между приходом и расходом продукции – явление достаточно редкое и наблюдается в наиболее стабильных сообществах, например, в тропической зоне. Однако это создает объективные трудности для развития там сельского хозяйства.
Человек, выжигая пышный тропический лес надеется получить на освободившейся территории высокие урожаи. Однако вскоре оказывается, что почвы на этой территории абсолютно бесплодны – вся годовая продукция росшего на этом месте леса потреблялась различными консументами и редуцентами и в почвах ничего не откладывалось.
Кроме первичной и вторичной продукции биогеоценозов, различают промежуточную и конечную продукцию.
Промежуточная продукция - это продукция, которая после потребления членами биогеоценоза снова возвращается в круговорот веществ этой системы.
Конечная продукция - это продукция, которая выводится за пределы данной экосистемы.
Например, продукция, получаемая человеком в процессе возделывания сельскохозяйственных культур, разведения домашних животных, охоты, промысла и т.д.
Продуктивность различных экосистем неодинакова и зависит от ряда экологических факторов, в первую очередь, климатических (тепло, влага и др).
При этом первичная продукция органического вещества в экосистемах, богатых жизнью, может превосходить продукцию сравнительно бедных экосистем более, чем в 50 раз.
Наиболее продуктивны экосистемы эстуариев и коралловых рифов (средняя продуктивность достигает 20 г / м2 в сутки), влажных тропических лесов и болот (средняя продуктивность составляет 10 г / м2 в сутки).
Высокопродуктивные экосистемы встречаются там, где климатические условия благоприятны, особенно при дополнительном поступлении в экосистему энергии извне.
Поступление энергии со стороны абиотических компонентов уменьшает затраты живых организмов на поддержание собственной жизнедеятельности, т.е. они компенсируют свои затраты на дыхание.
Например, энергия приливов повышает продуктивность природной прибрежной экосистемы, компенсируя потери энергии при дыхании.
Низкой продуктивностью (0,1-0,5 г / м2 в сутки) характеризуются экосистемы пустынь и тундр, в которых дефицит влаги и тепла лимитирует развитие низшего трофического уровня, а также открытые воды морей и океанов, где при избытке воды объем органических веществ сравнительно невысок.
При этом необходимо отметить, что большая часть земного шара покрыта океанами и пустынями с низкой продуктивностью, тогда как высокая продуктивность характерна для сравнительно незначительных участков Земли (эстуарии, коралловые рифы, болота, влажные леса).
Изменение первичной продуктивности экосистем по направлению с севера на юг происходит в следующем порядке:
в наземных арктических биогеоценозах продуктивность низкая, а арктические моря, так же как и антарктические являются высокопродуктивными;
в тропиках огромная часть суши занята малопродуктивными пустынями, бедны и моря этой зоны;
в экваториальной зоне располагаются самые высокопродуктивные биогеоценозы коралловых рифов, эстуариев, болот и особенно влажных тропических лесов.
По мере продвижения с севера на юг увеличивается удельное количество солнечной энергии, попадающей на единицу поверхности Земли, что приводит к большему количеству видов, накоплению более значительной биомассы и повышению продуктивности экосистем суши.
В морских экосистемах иная ситуация, чем на суше.
Высока продуктивность северных морей, а также морей крайних южных широт, где из глубин поступают холодные воды, богатые кислородом и биогенами. В теплой воде кислород растворяется хуже и мало биогенов (тропики богаты видами, но сравнительно мало продуктивны).
Общая чистая первичная продуктивность на Земле составляет 170 млрд. тонн в год, из которых 115 млрд. тонн дают экосистемы суши, а 55 млрд. тонн - экосистемы морей.
Вторичная продукция (биомасса гетеротрофных организмов, прежде всего животных - зоомасса), во много раз меньше первичной продукции (биомассы растений - фитомассы).
В разных экосистемах зоомасса составляет незначительную долю биомассы (от 0,05% до 5% всей биомассы), тем не менее животные суши играют большую роль в регулировании процессов, происходящих в отдельных экосистемах и биосфере в целом.
Совершенно очевидно, что жизнь людей, их производственная деятельность зависят от продуктивности основных биогеоценозов, от первичной продукции и ее мирового распределения.
Питание людей обеспечивается главным образом сельскохозяйственными культурами, занимающими около 10% площади суши и дающие примерно 9,1 млрд. тонн органического вещества в год, что составляет значительную часть мировых ресурсов.
Кроме того, огромная масса первичной продукции используется человеком как техническое сырье в промышленности и быту (топливо, хлопок, лен, эфиромасличные культуры и др.), причем около 50% теряется в отходах.
Но человек потребляет не только первичную продукцию. Он изымает из биосферы большое количество вторичной продукции в виде животной пищи, расходы которой подсчитать очень трудно.
Таким образом, имеющиеся представления о продуктивности экосистем и мировом распределении первичной продукции дают возможность ориентироваться в обстановке, сложившейся на нашей планете и на строго научной основе разрабатывать мероприятия по рациональному использованию природных ресурсов.
3.5. Классификация экосистем по энергетическому признаку
По энергетическому признаку выделяют четыре фундаментальных вида экосистем:
1) Экосистемы, движимые солнцем и несубсидируемые другими естественными источниками, т.е. природные системы, полностью зависящие от прямого солнечного излучения.
К таким экосистемам относятся:
- открытые участки океанов;
- большие глубокие озера;
- крупные участки горных лесов.
Экосистемы такого типа получают мало энергии и имеют малую продуктивность. Однако они имеют очень важное значение, так как занимают огромные площади и здесь очищаются большие объемы воздуха, возвращается в оборот вода, формируются климатические условия и т.д.
2) Экосистемы, движимые солнцем и субсидируемые другими естественными источниками (дополнительная энергия в виде дождя, ветра, органических веществ, минеральных элементов и т.д.).
Примерами таких экосистем являются: эстуарии рек, морские проливы и лагуны.
Приливы и течения способствуют более быстрому круговороту минеральных элементов питания и поэтому эстуарии более плодородны, чем прилегающие участки океана или суши.
3) Экосистемы, движимые солнцем и субсидируемые человеком.
Примером их являются агроэкосистемы.
Дополнительная энергия – в виде горючего, органических и минеральных удобрений, пестицидов, стимуляторов роста и т.д.
4) Экосистема, движимая топливом, - индустриально-городская экосистема, в которой энергия топлива не дополняет, а заменяет солнечную энергию.
Потребность в энергии плотно заселенных городов на 2-3 порядка больше того потока энергии, который поддерживает жизнь в естественных экосистемах, движимых солнцем.
4. Динамика и стабильность экосистем
Любая экосистема, приспосабливаясь к изменениям внешней среды, находится в состоянии динамики.
Динамика экосистем выражается в различных масштабах времени: изменения в биоценозах могут иметь суточную или сезонную динамику, длиться на протяжении ряда лет или же охватывать целые геологические эпохи, отражая развитие биосферы в целом.
Различают динамику циклическую и направленную.
При циклической динамике происходит суточное и сезонное изменение активности жизнедеятельности организмов, или периодическое изменение численности отдельных видов в многолетнем ряду.
Направленная динамика представляет собой поступательное развитие экосистем, которое приводит либо к внедрению в экосистемы новых видов, либо смена одних видов другими, что в конечном итоге приводит к смене биоценоза и экосистемы в целом.
4.1. Суточная и сезонная динамика экосистем.
Суточная динамика экосистем
В каждом биоценозе имеются группы организмов, активность жизни у которых приходится на разное время суток. Одни виды организмов активны в ночное время, а днем скрываются в убежищах. Другие виды животных отличаются дневной активностью и пассивны ночью.
При этом суточную динамику биоценозов обеспечивают не только животные, но и растения. У растений в течение суток также изменяется интенсивность и характер физиологических процессов – ночью не происходит фотосинтез, нередко у растений цветки раскрываются только в ночные часы и опыляются ночными животными, большинство видов растений приспособлены к опылению днем.
Суточная динамика в биоценозах, как правило, выражена тем сильнее, чем значительнее разница температур, влажности и других факторов среды днем и ночью.
В результате в составе и в соотношении отдельных видов биоценоза той или иной экосистемы происходят периодические изменения, так как отдельные организмы на определенное время выключаются из него.
Разделение периодов активности во времени снижает уровень конкуренции и открывает возможность для сосуществования видов со сходными биологическими требованиями.
В целом расхождение в суточной активности приводит к усложнению биоценоза, повышению биологического разнообразия и более полному использованию ресурсов среды.
Большое значение в суточной динамике биоценозов имеют суточные миграции живых организмов, которые зависят от изменения характера действия различных факторов в течение суток.
Особенно типичны вертикальные суточные миграции морского планктона.
- представители зоо- и фитопланктона в морях днем держатся на глубине от 100 до 350 м, а ночью поднимаются в поверхностные слои.
Вертикальные суточные миграции свойственны и почвенным обитателям.
Таким образом, суточная динамика экосистем связана главным образом с ритмикой природных явлений и носит строго периодический характер.
Кроме того, могу происходить и непериодические изменения активности и численности тех или иных компонентов экосистемы в течение суток, связанные с действием нерегулярных факторов среды.
Например, сильные дожди или засухи приводят к перемещению животных, изменению их активности, влияют на интенсивность некоторых жизненных процессов у растений:
- во время сильных ливней морской планктон из-за опреснения поверхностных слоев воды мигрирует в глубину;
- в летний период, когда суточные изменения температуры наиболее экстремальны, ряд дневных видов меняет характер активности на ночной (некоторые насекомые, змеи).
Сезонная динамика экосистем
Более значительные отклонения в биоценозах наблюдаются при сезонной динамике. Эти отклонения обусловлены биологическими циклами организмов, которые зависят от сезонной цикличности природных явлений.
Например:
Смена времен года оказывает значительное влияние на жизнедеятельность животных и растений:
- у животных – спячка, зимний сон, диапауза и миграции;
- у растений – периоды цветения, плодоношения, активного роста, листопада и зимнего покоя.
Сезонной изменчивости подвержена нередко и ярусная структура биоценоза. Отдельные ярусы растений в соответствующие сезоны года могут полностью исчезать (например, травянистый ярус).
Длительность биологических сезонов в разных широтах неодинакова. В связи с этим сезонная динамика биоценозов арктической, умеренной и тропической зон различна.
Наиболее четко сезонная динамика выражена в экосистемах умеренного климата и в северных широтах, которые отличаются резкими изменениями физических параметров среды летом и зимой.
Например:
- в тундрах, для которых в летнее время характерен большой набор видов птиц, насекомых и других животных; а на зиму большая часть из них мигрирует на юг (большинство птиц, северные олени), а другие впадают в оцепенение и выключаются из активной жизни (насекомые, некоторые беспозвоночные животные).
- длинная полярная ночь закрывает возможность фотосинтеза для растений в тундровых экосистемах.
В тропиках сезонная динамика выражена не столь четко, но все же имеет место.
Наиболее распространенная ее форма – смена засушливых и влажных периодов - явление не столь ритмичное, как смена летних и зимних периодов, но биологически не менее значимое.
Во всех случаях уменьшение числа активных видов влечет за собой снижение общего уровня биогенного круговорота веществ и является важным фактором регуляции биосферных процессов.
В связи с тем, что характер суточных и сезонных изменений более или менее постоянен в течение длительного времени (столетия или даже тысячелетия), в биоценозах исторически сформировались механизмы, приводящие сообщество в целом в соответствие с периодичностью изменения условий обитания.
Поэтому, характерной чертой процесса суточной и сезонной динамики является сохранение принципиальных свойств данной экосистемы, поддержание ее целостности и функциональной устойчивости.
Биоценоз испытывает лишь периодические колебания качественных и количественных характеристик.
4.2. Экологические сукцессии
Динамика биоценозов может быть принципиально иной, когда под влиянием комплекса факторов меняются фундаментальные свойства экосистем и происходит их изменение в виде серии сменяющихся друг друга сообществ.
Экологическая сукцессия (от лат. преемственность, наследование) представляет собой развитие биоценозов, при котором происходит последовательная смена во времени одного сообщества другим.
Последовательный ряд постепенно и закономерно сменяющих друг друга в сукцессии сообществ называется сукцессионной серией (сукцессионным рядом).
Причинами подобных смен могут являться внешние по отношению к биоценозу факторы, действующие длительное время в одном направлении.
Такими факторами могут быть как естественные (отступление ледников, наводнения, землетрясения, извержения вулканов, пожары и т.д.,), так и антропогенные (расчистка лесных угодий, распашка степей, добыча полезных ископаемых, создание прудов и водохранилищ, пожары, загрязнение экосистем и т.д.)
Сукцессии, обусловленные действием внешних факторов, называются экзогенетическими.
Эндогенетические сукцессии происходят в результате процессов, которые происходят внутри самого биоценоза.
Эндогенетические сукцессии наблюдаются в природе, когда в процессе своего развития сообщество изменяет среду так, что она становится более благоприятной для другого сообщества. Формирующееся новое сообщество в свою очередь делает среду еще более неблагоприятной для прежнего сообщества. Менее приспособленные к новым условиям среды виды замещаются более приспособленными и так до тех пор, пока не появятся виды, которых условия среды полностью устраивают и они уже не замещаются другими видами.
Сукцессия заканчивается формированием конечного, завершающего или так называемого климаксного сообщества, наиболее адаптированного по отношению к условиям среды, способного поддерживать себя неограниченно долго, внутренние компоненты которого уравновешены друг с другом и окружающей средой.
Иными словами, климаксный биоценоз находится в состоянии гомеостаза.
Однако существование абсолютно климаксного биоценоза невозможно, так как взаимное воздействие его компонентов никогда не прекращается и в результате совершаются сукцессии. Этот процесс постоянен, так как развитие биоценозов, как и любого природного явления, не может остановиться.
Со временем микроорганизмы, растений и животные в биоценозе приобретают лишь некоторую относительную устойчивость – сбалансированность отдельных компонентов в сообществе. Но такое климаксное состояние биоценоза не может быть постоянным во времени и пространстве.
Поэтому, термин «климакс» можно использовать лишь условно в смысле приобретения биоценозом определенной замедленности в развитии, поскольку даже при кажущейся неизменности в биоценозе все-таки протекают процессы, подготавливающие его к сукцессии.
Процессы сукцессии в природе могут занимать различные временные промежутки.
Известны изменения экосистем, идущие на протяжении многих столетий, которые охватывают целые геологические периоды и отражают общие пути эволюции биосферы. Такие смены экосистем называются вековыми сукцессиями.
Процессы сукцессии, идущие в геологических масштабах времени, тесно связаны с геологическими и климатическими изменениями, а также с эволюцией видов.
Если в результате изменения условий среды одни виды вымирают, а другие изменяются под действием естественного отбора, данный процесс рассматривается как эволюционная сукцессия.
Примерами вековой смены экосистем могут служить:
изменения сообществ растений и животных по мере отступления ледников после крупных оледенений;
сукцессия от широколиственных лесов к пустыням в северной части Средней Азии, связаны с вековым изменением климата.
В большинстве случаев процессы сукцессии совершаются в сравнительно короткие промежутки времени, измеряемые годами и десятилетиями.
Например:
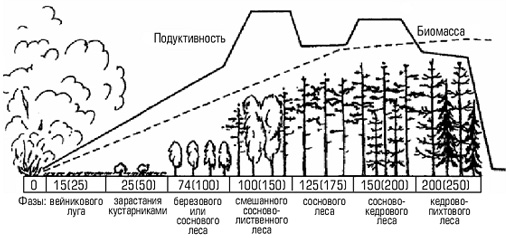
лесной пожар может моментально уничтожить сложившийся на протяжении тысячелетий устойчивый биоценоз и на пожарище начинает довольно быстро создаваться новое сообщество. При этом будет наблюдаться частая смена ряда сообществ. В конечном итоге восстанавливается биоценоз исходного типа или же он незначительно отличается от первоначального;
зарастание небольшого озера с последующим появлением на его месте болота, а затем леса. Вначале по краям озера образуется сплавна – плавающий ковер из осок, мхов и других растений. Постоянно озеро заполняется отмершими остатками растений – торфом. Образуется болото, постепенно зарастающее лесом.
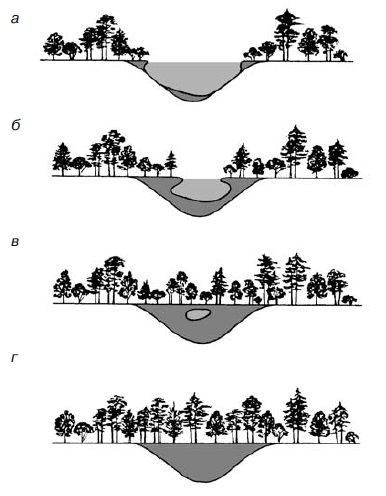
Рис. 1. Процесс исчезновения озера: а — озеро; б — по мере увеличения в озере осадочных материалов оно мелеет; в — в дальнейшем озеро превращается в болото и покрывается растительностью; г— в конце концов это место превращается в сушу. Обычно этот процесс занимает несколько тысяч лет даже для мелководных озер
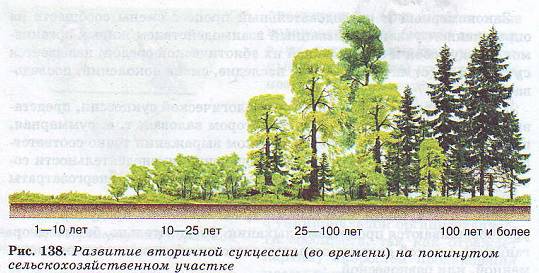
Формирование елового леса на заброшенных пашнях – другой классический пример сукцессии.
Еловый лес в своем развитии проходит несколько этапов.
Первыми на заброшенных пашнях появляются береза, осина и ольха, поскольку семена этих деревьев легко разносятся ветром. Такие первопоселенцы обычно называют пионерами. Наиболее стойкие из них заселяют заброшенную или распаханную территорию, утверждаются там и постепенно изменяют среду, создавая тем самым новые условия, к которым сами со временем оказываются неприспособленными.
Эти условия становятся пригодными для растений-захватчиков, которые вытесняют пионеров и начинают доминировать в сообществе до тех пор, пока в результате их деятельности вновь не изменяться условия и они не начнут замещаться более приспособленными формами.
Условия, благоприятные для появления ели, создаются только после смыкания крон берез, которое происходит примерно через 30-50 лет. Постепенно формируется смешанный лес. Однако он существует сравнительно недолго, так как светолюбивые березы не выносят затенения и под пологом елей их возобновления не происходит.
Устойчивый еловый лес на заброшенной пашне образуется примерно через 80—120 лет после первых всходов березы.
В процессе развития березового, а затем и елового леса в биоценоз включаются все новые виды растений и животных, которые сопровождают доминантов. Одновременно происходит замещение одних видов другими. По мере того как увеличивается их число, в сообществе возникают и заполняются новые экологические ниши.
Таким образом, приведенные примеры убедительно свидетельствуют о том, что экологическая сукцессия является результатом изменений, которые вносятся в среду обитания самими сообществами.
Это закономерный, направленный процесс, который заканчивается образованием климаксового сообщества, характеризующегося максимальной величиной биомассы и наибольшим разнообразием слагающих его видов.
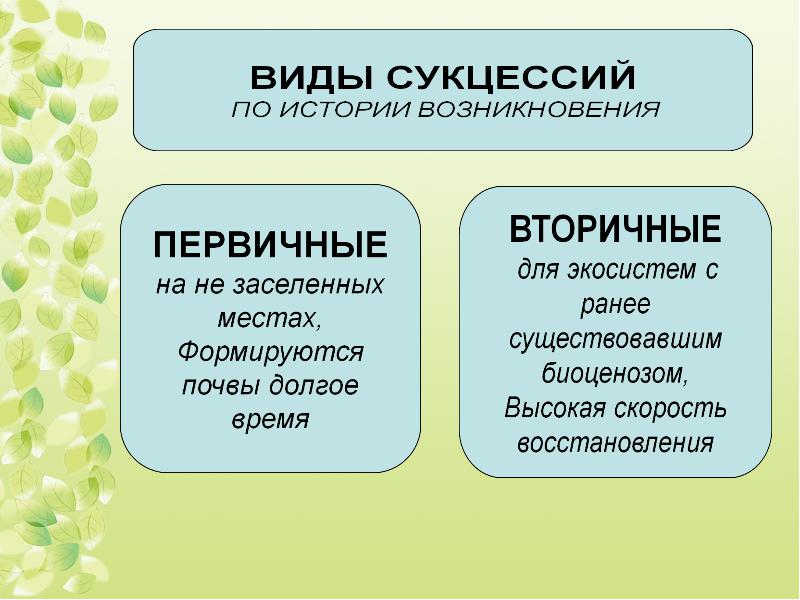
По общему характеру сукцессии подразделяются на первичные и вторичные.
Первичной называется сукцессия, которая начинается на абсолютно лишенном жизни месте.
При первичных сукцессия скорость изменения сообществ, как правило, невелика. Сукцессионные ряды, последовательно сменяющие друг друга, удерживаются значительный промежуток времени, а достижение экосистемой климаксового состояния затягивается иногда на многие столетия.
Примерами первичных сукцессий в масштабе геологических периодов являются формирование фауны и флоры на скалах, песках, лавовом потоке, в послеледниковый период, после поднятия островов в океане.
Сукцессия называется вторичной, если сообщество развивается на месте, где раньше существовал хорошо развитый биоценоз.
В таких местах обычно сохраняются богатые жизненные ресурсы. Поэтому вторичные сукцессии приводят к образованию климаксового сообщества значительно быстрее, чем первичные.
В современных условиях вторичные сукцессии наблюдаются повсеместно. Они обусловлены последствиями, возникающими в результате пожаров, наводнений, распашки степей, вырубки лесов, осушения болот.
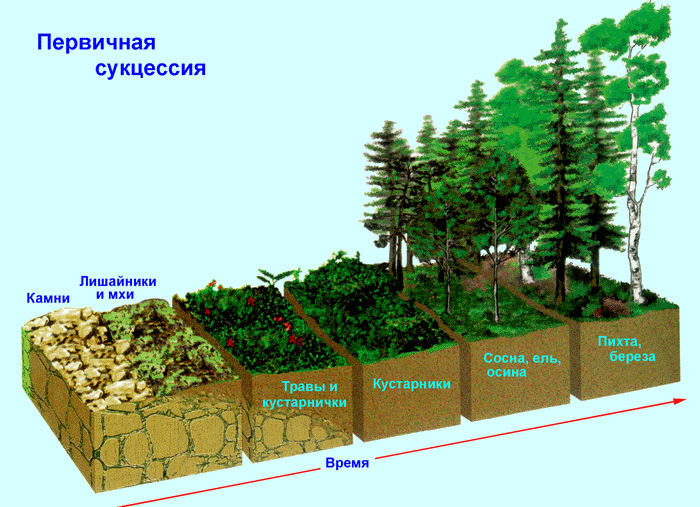
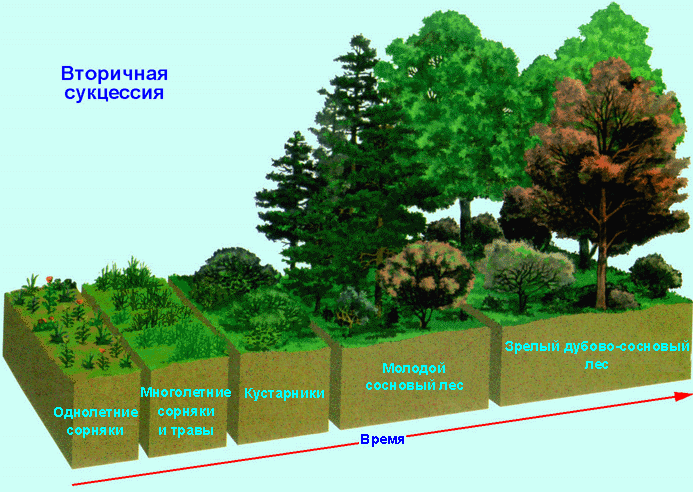
Для любой сукцессии, особенно первичной, характерны следующие общие закономерности протекания процесса.
Видовой состав растений и животных в процессе сукцессии непрерывно меняется.
Это свидетельствует о том, что виды, доминирующие на начальных стадиях сукцессии, по мере достижения биоценозом климаксного состояния постепенно утрачивают это свойство.
Такое явление наглядно прослеживается на рассмотренном выше примере с заменой березняка еловым лесом.
Биомасса органического вещества увеличивается по ходу сукцессии.
Совершенно очевидно, что с течением времени как в наземной, так и в водной среде увеличивается общая биомасса сообщества и происходит накопление мертвого органического вещества.
Видовое разнообразие повышается по ходу сукцессии.
Наиболее интенсивно увеличивается видовое разнообразие гетеротрофных организмов.
Видовое разнообразие автотрофов увеличивается на сравнительно ранних этапах сукцессии, тем самым обеспечивая максимум первичной продукции и создавая благоприятные условия для увеличения видового разнообразия гетеротрофов, которое растет до относительно поздней стадии сукцессии.
Снижение чистой продукции сообщества и соответственное повышение интенсивности дыхания – самые важные явления сукцессии.
Это объясняется тем, что продуктивность сообщества на разных стадиях сукцессии различна:
- на начальных стадиях приход продуктивности (создание органического энергетического материала) превышает ее расход (затраты на дыхание). В результате быстро увеличивается биомасса сообщества;
- по мере приближения сукцессии к климаксному состоянию продуктивность, достигнув максимума, начинает уменьшаться;
- в климаксном биоценозе продуктивность и дыхание (приход и расход) уравновешиваются и увеличения как первого, так и второго процессов не наблюдается: биоценоз достигает более или менее стабильного, гомеостатического состояния.
Это означает, что в климаксном биоценозе все создаваемые пищевые вещества используются, вследствие чего чистая продукция остается невысокой, а количество биомассы становится максимальным.
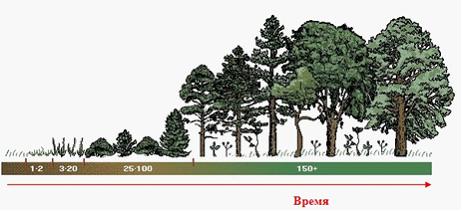
4.3. Стабильные и нестабильные экосистемы.
К антропогенным экосистемам относятся агроэкосистемы и урбоэкосистемы.
Агроэкосистемы (агроценозы) – это искусственные экосистемы, возникающие в результате сельскохозяйственной деятельности человека (пашни, сенокосы, пастбища, сады, парки, огороды, лесные насаждения и т.д.).
Агроэкосистемы создаются человеком для получения высокой чистой продукции автотрофов (урожай).
В них, так же, как в естественных сообществах, имеются пищевые цепи, состоящие из:
- продуцентов (культурные растения и сорняки);
- консументов (насекомые, птицы, грызуны и т.д.)
- редуцентов (грибы и бактерии).
Однако обязательным звеном пищевых цепей в агроэкосистемах является человек, который своим трудом создает данный агроценоз и обеспечивает его высокую продуктивность, а затем собирает и использует урожай.
В структуре и энергетике естественных экосистем и агроэкосистем существует ряд различий:
1. Способность к саморегулированию является одним из самых существенных различий между естественными экосистемами и агроэкосистемами.
Важнейшей особенностью любой естественной экосистемы является способность к саморегулированию, т.е. удерживанию своих основных параметров во времени и пространстве.
Агроценозы в отличие от природных экосистем не являются саморегулирующими системами, а регулируются человеком.
Задачей такой регуляции является повышение продуктивности агроценоза. Для этого орошаются засушливые и осушаются переувлажненные земли, уничтожаются сорняки и поедающие урожай вредители, меняются сорта культивируемых растений и вносятся удобрения.
2. Естественные экосистемы, как правило, более стабильны и обладают большим видовым разнообразием, чем агроценозы.
Причем чем больше видовое разнообразие, тем стабильнее экосистема. В таких экосистемах формируются сложные пищевые взаимоотношения, сложные цепи питания. В случае резкого увеличения численности популяции какого-либо вида немедленно вступают в действие механизмы, регулирующие ее.
Несколько по иному проявляется стабильность в агроценозах.
Агроценозы отличаются от природных экосистем незначительным количеством видов, но высокой их численностью; при этом численность этих видов постоянно поддерживается и регулируется человеком.
Например, отдельные виды животных, не имея биологических конкурентов, могут интенсивно размножаться и становятся вредителями, угрожающими существованию агроценоза. Поэтому для его сохранения требуется вмешательство человека с использованием мер для уничтожения этих видов животных.
Кроме того, человек, собирая урожай, изымает из агроценозов большую часть продукции, которая из-за этого не поступает в детритные цепи питания. В результате этого почва обедняется минеральными веществами, необходимыми для жизнедеятельности растений. Поэтому для сохранения агроценоза необходимо постоянно вносить в почку удобрения.
Экологическая устойчивость агроценозов невелика. Без участия человека агроценозы зерновых и овощных культур существуют не более года, многолетних трав – 3-4 года, а плодовых культур -20-30 лет. Затем такие агроценозы распадаются или отмирают.
3. В природных экосистемах действует естественный отбор, отметающий не конкурентоспособные формы организмов, тем самым обеспечивая устойчивость экосистемы.
В агроценозах действует преимущественно искусственный отбор, направленный на максимальную продуктивность растений и животных, необходимых человеку, а не тех, которые лучше приспособлены к окружающим условиям среды.
4. Различия в использовании энергии.
Для природных экосистем единственным источником энергии является солнечная энергия.
Агроценозы помимо солнечной энергии получают дополнительную энергию от человека: производство удобрений, препаратов против сорняков и вредителей, болезней, орошение или осушение полей. Без такой дополнительной энергии существование агроценозов практически невозможно.
5. Баланс питательных элементов.
В природных экосистемах первичная продукция растений потребляется в многочисленных цепях питания и вновь возвращается в систему биологического круговорота в виде диоксида углерода, воды и минеральных веществ.
В агроценозе совершается неполный круговорот веществ, поскольку значительную их часть человек изымает в виде урожая. Поэтому для возмещения потерь необходимо постоянно вносить в почву удобрения.
Урбоэкосистемы – это искусственные экосистемы, возникающие в результате развития городов, и представляющие собой сосредоточие населения, жилых зданий, промышленных, бытовых, культурных объектов и т.д.
В составе урбоэкосистем можно выделить следующие территории:
промышленные зоны, где сосредоточены промышленные объекты различных отраслей хозяйства и являющиеся основными источниками загрязнения окружающей среды;
селитебные зоны – жилые или спальные районы с жилыми домами, административными зданиями, объектами быта, культуры, просвещения и т.д.;
рекреационные зоны, предназначенные для отдыха людей (городские парки, базы отдыха и т.д.);
транспортные системы и сооружения, пронизывающие всю городскую систему (автомобильные и железные дороги, метрополитен, заправочные станции, гаражи, аэродромы и т.д.).
Существование урбоэкосистем поддерживается за счет агроэкосистем и энергии горючих ископаемых и атомной промышленности.
Основные типы экосистем
5.1. Наземные экосистемы
Наземные экосистемы:
1. Тундра: арктическая и альпийская;
2. Бореальные хвойные леса;
3. Листопадный лес умеренной зоны;
4. Степь умеренной зоны;
5. Тропические злаковники и саванна;
6. Чапараль (районы с дождливой зимой и засушливым летом);
7. Пустыня: травянистая и кустарниковая;
8. Полувечнозеленый тропический лес (районы с выраженным влажным и сухим сезонами);
9. Вечнозеленый тропический дождевой лес.
Характеристика наземных экосистем
Размещение по земной поверхности основных наземных биомов определяют два абиотических фактора - температура и количество осадков. Климат в разных районах земного шара неодинаков. Годовая сумма осадков меняется от 0до 2500мм и более. При этом они выпадают равномерно в течении года или их основная доля приходится на определенный период - влажный сезон. Среднегодовая температура также варьируется от отрицательных величин до 38° C. Температуры могут быть практически постоянными в течении всего года (у экватора) или меняться постоянно.
Тундры (в северном полушарии к северу от тайги). Климат очень холодный с полярным днем и ночью, среднегодовая температура ниже 0° C. За несколько недель короткого лета земля оттаивает не более чем на один метр в глубину. Осадков менее 200-300 мм в год. Растительность: отсутствуют деревья, господствуют медленно растущие лишайники, мхи, травы (злаки и осоки), стелющиеся или карликовые кустарнички (брусника, черника) и кустарники (карликовая береза). Животный мир небогат, встречаются крупные травоядные копытные - северный олень (Евразия) и карибу (Северная Америка), мелкие роющие млекопитающие (лемминги), хищники (песец, горностай, ласка). Среди птиц преобладают полярная сова, ржанка, пуночка. Среди насекомых обильны двукрылые. Почвы тундровые - бедные с малой мощностью над слоем вечной мерзлоты. Очень ранимые экосистемы из-за медленного их восстановления.
Бореальные хвойные леса (тайга) (северные районы Евразии и Северной Америки). Климат: долгая и холодная зима, много осадков выпадает в виде снега. Растительность: господствуют вечнозеленые хвойные леса (ель, пихта, сибирская кедровая сосна, сосна обыкновенная, лиственницы) с мощной лесной подстилкой. Животный мир: крупные травоядные копытные (лось), мелкие растительноядные млекопитающие (барсук, белка, бурундук), хищники (медведь, рысь, росомаха, соболь, лисица, волк, норка), обилен гнус. Характерно множество болот и озер. Почвы подзолистые, дерново-подзолистые, мерзлотно-таежные - маломощные и бедные.
Листопадные леса умеренной зоны (широколиственные леса)
(Западная Европа, Восточная Азия, восток США).Климат сезонный с зимними температурами ниже ° C, осадков 750 - 1500 мм в год. Растительность: господствуют широколиственные листопадные породы деревьев (дуб, бук, клен, липа, ясень, граб), кустарниковый подлесок, мощная лесная подстилка. Животный мир: Млекопитающие (лоси, медведи, рыси, лисицы, волки, белки, землеройки), птицы (дятлы, дрозды, совы, соколы). Биота адаптирована к сезонному климату: спячка, миграции, состояние покоя в зимние месяцы. Почвы бурые и серые лесные. В этих районах человеческая цивилизация получила наибольшее развитие, поэтому большая часть широколиственных лесов заменена культурными сообществами.
Степи умеренной зоны (в Евразии) и их аналоги: прерии (в Северной Америки), пампасы (в Южной Америки), туссоки (в Новой Зеландии). Климат сезонный, лето от умеренного теплого до жаркого, зимние температуры ниже ° C, осадков 250-750 мм в год. Растительность: господствуют дерновинные злаки. Животный мир: крупные растительноядные млекопитающие - бизоны, вилорогие антилопы, сайгаки и др.; мелкие роющие млекопитающие (суслики, сурки, кролики, полевки), хищники (волки, львы, гиены и др.), разнообразные птицы. Почвы: черноземы и каштановые. Большая часть степей используется в настоящее время под пашню, пастбища, сенокосы и т. д.
Чапарраль (Средиземноморье, южный берег Австралии, в Калифорнии, Мексике и Грузии). Климат мягкий умеренный, осадков 500-700 мм. Растительность: деревья и кустарники с жесткими вечнозелеными листьями (лавр, дуб, дикая фисташка и др.) Почвы коричневые и серо-коричневые.
Тропический грасленд и саванны (Центральная и Восточная Африка, Южная Америка, Австралия, значительная часть Южной Индии). Климат сухой и жаркий большую часть года, температура высокая круглый год, осадков 250-750 мм в год, распределяются неравномерно по сезонам. Растительность: травянистая растительность (злаковые) с редкими листопадными деревьями (баобабы, акации, пальмы). Животный мир: крупные растительноядные млекопитающие (антилопы, зебры, жирафы, носороги, слоны), хищники (львы, леопарды, гепарды), птицы (африканский страус, грифы). Много кровососущих насекомых, например, муха цеце. Почвы красные ферраллитные, красно-бурые и коричнево-красные. На распаханных землях выращивают злаковые, хлопчатник, арахис, сахарный тростник.
Пустыни травянистая и кустарниковая (некоторые районы Африки, Большой Бассейн и юго-запад США, север Мексики и др.). Климат очень сухой, с жарким днем и холодными ночами, осадков менее 200-250 мм в год. Растительность: ксерофитные травы и редкостойный кустарник, кактусы. Корневые системы у растений обширные, поверхностные, перехватывающие влагу редких осадков или стержневые корни, проникающие в землю до уровня грунтовых вод (30 м и глубже). Животный мир: разнообразные грызуны (тушканчики, суслики), копытные (вилорогая антилопа и др.), хищники (волк, койот и др.) Из птиц саджа, рябки, жаворонки. Почвы светло-бурые, сероземы, такыры. Экосистемы хрупкие, легко нарушаются в результате перевыпаса, ветровой и водной эрозии.
Полувечнозеленые сезонные листопадные тропические леса (тропическая часть Азии, Центральная Америка). Климат со сменой сухого (4-6 месяцев)и влажного сезонов, среднегодовое количество осадков 800-1300 мм в год. Растительность: господствуют леса. Животный мир: практически также богат, как в вечнозеленых тропических дождевых лесах. Характерны слоны, жирафы, буйволы. Почвы красные ферраллитные.
Вечнозеленые тропические дождевые леса (север Южной Америки, Центральная Америка, западная и центральная часть экваториальной Африки, Юго-Восточная Азия, прибрежные районы северо-запада Австралии, острова Индийского и Тихого океанов). Климат без смены сезонов в связи с близостью к экватору, среднегодовая температура выше 17° C, среднегодовое количество осадков превышает 2000-2500 мм в год. Растительность. Деревья разной высоты образуют густой полог из многих ярусов (10-12 ярусов). Видовое разнообразие растений огромно. Животный мир видовой состав богаче, чем во всех биомах вместе взятых: Млекопитающие (обезьяны, ленивцы, ягуары), птицы (попугаи, колибри, туканы). Почвы красно-желтые ферраллитные - маломощные и бедные органическим веществом и минеральными элементами питания растений.
5.2. Водные экосистемы
Пресноводные экосистемы:
1. Лентические (стоячие воды): озера, пруды, водохранилища и др.;
2. Лотические (текучие воды): реки, ручьи, родники и др.;
3. Заболоченные угодья: болота, болотистые леса, марши (приморские луга).
Морские экосистемы:
1. Открытый океан (пелагическая экосистема);
2. Воды континентального шельфа (прибрежные воды);
3. Районы апвеллинга (плодородные районы с продуктивным рыболовством);
4. Эстуарии (прибрежные бухты, проливы, устья рек, лиманы, соленые марши и др.);
5. Глубоководные рифтовые зоны.
Помимо основных типов природных экосистем (биомов)различают переходные типы - экотоны. Например, лесотундра, смешанные леса умеренной зоны, лесостепь, полупустыни и др.
Характеристика водных экосистем
Распределение организмов в экосистемах зависит от степени освещенности. Выделяют следующие зоны: литоральная зона (толща воды, где солнечный свет доходит до дна), лимническая зона (толща воды до глубины, куда проникает всего 1% от солнечного света и где затухает фотосинтез), эвфотическая зона (вся толща воды - включает литоральную и лимническую зоны), профундальная зона (дно и толща воды, куда не проникает солнечный свет). В приточных водоемах выделяют перекаты (мелководные участки с быстрым течением: дно без ила, встречаются преимущественно прикрепленные формы перифитона и бентоса) и плесы (глубоководные участки: течение медленное, на дне мягкий илистый субстрат и роющие животные).
Лентические экосистемы (озера, пруды, водохранилища и др.).
Литоральная зона населена двумя группами растений: укрепившиеся на дне (камыши, рогозы, кувшинки, прикрепленные водоросли и др.) и плавающие (водоросли, рдесты и др.). Животные в литорали разнообразны, чем в других зонах водоема. Встречаются моллюски, коловратки, мшанки, личинки насекомых и др. Рыбы большую часть жизни проводят в литорали и здесь же размножаются. Многие обитающие здесь животные дышат кислородом атмосферного воздуха (лягушки, саламандры, черепахи и др.). Зоопланктон представлен ракообразными, имеющими большое значение для питания рыб (дафнии и др.). Лимническая зона. Продуценты представлены фитопланктоном. В водоемах умеренного пояса "цветение" весной связано с массовым развитием диатомовых, летом - зеленых, осенью - азотфиксирующих сине-зеленых водорослей. Зоопланктон представлен растительноядными ракообразными и коловратками. Нектон лимнической зоны - только рыбы. Профундальная зона около дна представлена бентосными формами - личинками насекомых, моллюсками, кольчатыми червями, сапротрофными бактериями и грибами.
Лотические экосистемы (реки, родники, ручьи и др.) отличаются от стоячих водоемов следующими особенностями: 1) наличие течения; 2) более активный обмен между водой и сушей; 3) более высокое содержание кислорода и более равномерное распределение; 4) преобладание детритных цепей питания. Выделяют лотические сообщества перекатов и плесов. На перекатах поселяются организмы, способные прикрепится к субстрату (например, нитчатые водоросли) или хорошие пловцы (например, форель). На участках плеса сообщества напоминают прудовые. В больших реках прослеживается продольная зональность: в верховьях - сообщества перекатов, в низовьях и дельте - плесов, между ними местами могут возникать и те и другие. Видовой состав рыб к низовьям объединяется, но увеличиваются их размеры.
Заболоченные участки и болота бывают низинные (имеют, как правило, питание подземными водами) и верховые (питаются атмосферными осадками). Верховые могут встречаться в любом понижении или даже на склонах гор, низинные возникают вследствие зарастания озер и речных стариц. Здесь распространены болотные растения. Болотные почвы и торфяники содержат много углерода. Их сельскохозяйственная отработка приводит к выделению в атмосферу большого количества углекислого газа.
Область бесконечного шельфа является самой богатой в фаунистическом отношении. Прибрежная зона очень благоприятна по условиям питания, даже в дождевых тропических лесах нет такого разнообразия жизни, как здесь.
Районы апвеллинга расположены вдоль западных пустынных берегов континентов. Здесь наблюдается апвеллинг - подъем холодных вод с глубины океана, так как ветры перемещают воду от крутого материкового склона, а взамен ей из глубины поднимается вода, обогащенная биогенными элементами. Эти районы богаты рыбой и птицами, живущими на островах.
Эстуарии, лиманы, устья рек, прибрежные бухты и т. д. - прибрежные водоемы, представляющие собой экотоны между пресноводными и морскими экосистемами. Это высокопродуктивные районы, где наблюдаются аутвеллинг - привнос биогенных элементов с суши. Они обычно входят в литоральную зону и подвержены приливам и отливам. Здесь встречаются болотные и морские травы, водоросли, рыба, крабы, креветки, устрицы и т. д.
Открытый океан беден биогенными элементами. Эти районы можно считать "пустынями" по сравнению с прибрежными водами. Арктические и антарктические зоны более продуктивны, так как плотность планктона растет при переходе от теплых морей к холодным, и фауна рыб и китообразных здесь значительно богаче. Продуцентом выступает фитопланктон, им питается зоопланктон, а тем в свою очередь нектон. Видовое разнообразие фауны снижается с глубиной. На глубине в стабильных местообитаниях сохранились виды из далеких геологических эпох.
Глубоководные рифтовые зоны океана находятся на глубине около 3000 м и более. Условия жизни в экосистемах глубоководных рифтовых зон очень своеобразны. Это полная темнота, огромное давление, пониженная температура воды, недостаток пищевых ресурсов, высокая концентрация сероводорода и ядовитых металлов, встречаются выходы горячих подземных вод, и т. д. В результате живущие здесь организмы претерпели следующие адаптации: редукция плавательного пузыря у рыб или заполнение его полости жировой тканью, атрофирование органов зрения, развитие органов светосвечения и др. Живые организмы представлены гигантскими червями (погонофорами), крупными двустворчатыми моллюсками, креветками, крабами и отдельными видами рыб. Продуцентами выступают сероводородные бактерии, живущие в симбиозе с моллюсками.
5.3. Различия водных и наземных экосистем.
Между водными и наземными экосистемами, несмотря на сходное проявление основных закономерностей, имеются и различия, обусловленные различными условиями внешней среды.
1. Структура водных экосистем значительно проще наземных.
Это объясняется тем, что во многих водных экосистемах, и прежде всего в глубоководных, нет атмосферы и почвы. Несколько сложнее структура мелководных экосистем, так как в прибрежных водных сообществах значительную роль играет и воздушная среда.
2. На организмы водных экосистем воздействуют такие факторы среды, которые не имеют никакого значения для организмов наземных экосистем, и наоборот.
При изучении наземных экосистем прежде всего приходится исследовать влажность и температуру среды, а для водных – температуру, газовый режим, растворенные и взвешенные в воде вещества.
3. Водные и наземные экосистемы характеризуются различным видовым составом растительных и животных организмов.
В водных экосистемах преобладают низшие растения, в первую очередь водоросли, тогда как в наземных – высшие цветковые растения.
Животное население водных экосистем представлено чрезвычайно широко, но высшие позвоночные животные имеют здесь второстепенное значение.
В наземных экосистемах, наоборот, высшие позвоночные, особенно млекопитающие и птицы, играют главную роль.
4. Водные и наземные экосистемы отличаются трофическими связями между организмами.
В водных экосистемах большая часть вещества и энергии включена в пастбищные цепи, а в наземных экосистемах наибольшее значение имеют детритные цепи.








Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!

