
Россия, Кабардино-Балкария.


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока
Была в сети 28.03.2017 13:46

Отарова Елена Хазреталиевна
учитель
54 года
Местоположение
Специализация
Мхи.Лекции для 6 класса.
Категория:
Биология
24.03.2016 23:09

Просмотр содержимого документа
«Мхи.Лекции для 6 класса.»
Занятие 6. Отдел Моховидные (Bryophyta)
Общая характеристика. Эти древнейшие высшие растения появились около 400 млн. лет назад (в девоне палеозойской эры). Они достаточно разнообразны (около 25 тыс. видов), распространены в увлажненных местообитаниях, но встречаются и в пустыне, и на бесплодной скальной поверхности (рис. 1).

Рис. 1. Разнообразие мохообразных.
Наличие многих примитивных черт в строении и жизнедеятельности позволяют рассматривать их в качестве боковой тупиковой ветви эволюции (рис. 2).
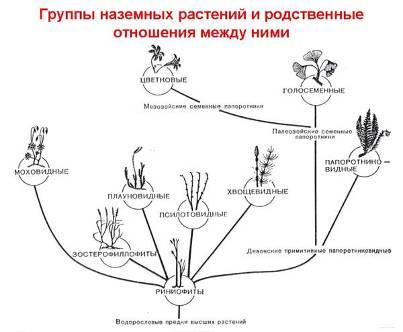
Рис. 2. Эволюция растительного мира.
1. Чередование поколений с преобладанием в цикле развития гаметофита. Гаметофит обычно многолетний. Он представляет собой листовидный таллом или растение в виде побега, расчлененного на стебель и листья (рис. 3). Спорофит, называемый у моховидных спорогоном, играет подчиненную роль. Он представляет собой цилиндрическую ножку, заканчивающуюся шаровидной, эллиптической или цилиндрической коробочкой, внутри которой образуется спорангий со спорами. Спорогон паразитирует на гаметофите, так как получает от него воду и необходимую пищу. Моховидные – единственная ветвь с регрессивным развитием спорофита.

Рис. 3. Строение мхов.
2. Вегетативное тело представлено либо слоевищем, либо слегка дифференцированным на стеблевую и листовую части. Настоящих корней и листьев у мхов нет. Функцию корней выполняют ризоиды – по большей части органы прикрепления, состоящие из вытянутых одиночных клеток у примитивных представителей (печеночников) или нитей из одного ряда клеток, разделенных косыми перегородками.
Листья являются выростами покровных тканей стебля, представляя собой пластинки из одного, реже – двух слоев клеток, с центральным тяжем, аналогичным жилке, и не имеют устьиц.
3. Поглощение и испарение воды у Моховидных осуществляется всей поверхностью гаметофита – очень древняя способность поглощать воду не по физиологическим, а физическим законам (капиллярность, гигроскопичность, набухание) (рис. 4).

Рис 4. Поглощение воды мхами (набухание).
4. У мхов нет настоящих проводящих элементов, только предшественники проводящих элементов: гидроиды (проводят воду) и липоиды (проводящие органические вещества) – длинные паренхимные клетки, выполняющие как запасающую, так и проводящую функции, нередко формирующие в центре стебля центральный проводящий пучок (рис. 5). Снаружи он окружен слоями крупных, тонкостенных, лишенных содержимого клеток, способных поглощать большое количество воды. Стебель покрыт корой из мелких, толстостенных клеток. Но бывают стебли, состоящие целиком из совершенно одинаковых клеток, без какой-либо дифференциации.

Рис. 5. Поперечные срезы стеблей листостебельных мхов. а – в центре стебля видны клетки центрального проводящего пучка, периферический ряд толстостенных клеток – наружная кора; б – центральный проводящий пучок отсутствует, внутреннее строение стебля довольно однообразно.
5. Фотосинтез Мохообразных характеризуется примитивными чертами: низкая продуктивность – в 40-50 раз меньше, чем у Цветковых, мелкие хлоропласты. В то же время фотосинтез у многих Мохообразных умеренных и холодных областей может протекать не только круглосуточно, но и круглогодично (рис. 6). Это примитивный признак – способность к анабиозу (при экстремальных условиях – дефиците влаги и больших перепадах температур – осуществляется низкий обмен веществ).

Рис. 6. Фотосинтез мхов при низких температурах.
6. Мейоз происходит не перед образованием гамет, а перед образованием спор. Гаметы образуются в результате митоза.
На зеленом гаметофите мохообразных из специальных клеток образуются половые органы – антеридии (мужские) и архегонии (женские). Антерозоиды (мужские половые клетки), созревая, выходят из антеридиев и устремляются хемотаксическим способом к архегониям с яйцеклетками. Происходит это только в капельно-жидкой среде – в воде, скапливающейся на поверхности почвы или в каплях дождя на листьях. Оплодотворенная яйцеклетка дает начало диплоидному спорогону, представляющему собой коробочку на ножке. В коробочке содержится специальная спорогенная ткань, из которой образуются споры. В процессе созревания споры проходят редукционное деление, поэтому они гаплоидны. К моменту созревания коробочки и спор крышечка, которой она накрыта сверху, отделяется, и споры высеваются. Из попавших в подходящие для прорастания условия спор вырастает протонема – «предросток» – представляющая собой в одних случаях тонкую нить из одного слоя клеток, очень напоминающую зеленую водоросль, в других – лопастное слоевище. На протонеме закладываются вегетативные почки, дающие начало взрослому гаметофиту.
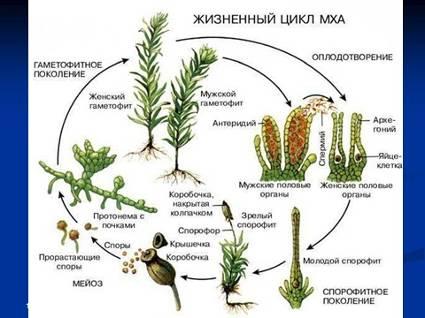
Рис. 7. Жизненный цикл мхов.
Вегетативно Мохообразные размножаются с помощью специальных органов (выводковых почек, листьев, частей листьев, веточек), вегетативно способен размножаться и спорофит (ножка).
7. Оплодотворение только при наличии воды – еще один примитивный признак (рис. 8).

Рис. 8. Оплодотворение у мхов в капельно-жидкой среде.
В основу классификации Мохообразных бриологи кладут морфологическое строение тела гаметофита, особенности строения ризоидов, строение и характер раскрывания коробочек, а также географическое распространение.
Класс Печеночники, или Печеночные мхи – Marchantiopsida
Класс объединяет около 6 (10) тыс. видов. Главная особенность Печеночников – дорзовентральное строение тела (рис. 9). Листья печеночников в отличие от листьев мхов всегда однослойные и состоят из однородных клеток. Ризоиды их только одноклеточные, хотя могут иметь разное строение.

Рис. 9. Маршанция (Marchantia polymorpha). Внешний вид.
Мужской и женский гаметофиты значительно отличаются внешне по форме развивающихся на них спорофитов (рис. 10).


Рис. 10. Талломы с мужскими (слева) и женскими (справа) спорогониями.
Для спорогония Печеночников характерно то, что в коробочке споры развиваются непосредственно под стенкой, т.е. она функционально является спорангием (рис. 11). Стенка при созревании либо сгнивает, либо растрескивается, но она не имеет механизмов для активного выбрасывания спор. Протонемы Печеночников очень слабо развиты. У слоевищных видов они по форме мало отличаются от взрослого гаметофита. У всех Печеночников обнаруживается микориза, отсутствующая у листостебельных мхов. Вегетативно размножаются – выводковыми почками.
Класс Листостебельные мхи –Bryopsida
Подкласс Настоящие мхи – Bryidae
Подкласс насчитывает около 14 тыс. видов. Среди них многие отличаются широким распространением (иногда в умеренных и холодных зонах обоих полушарий), высокой жизненной устойчивостью и большой фитоценотической ролью в растительном покрове Земли. Экологическое своеобразие и особое значение мхов в природных процессах во многом зависит от присущих им групповых форм роста, благодаря чему мхи могут на больших площадях создавать рыхлые или плотные покровы различной мощности (рис. 12). Тем самым мхи активно участвуют в формировании на поверхности континентов мощных влагоприемников в виде болот и замоховелых лесов, оказывающих существенное влияние на общую обеспеченность сушей влагой.

Рис. 11. Строение спорогониев маршанции. 1 – таллом с мужскими подставками; 2 – разрез через мужскую подставку; 3 – сперматозоиды; 4 – таллом с женскими подставками; 5 – продольный разрез молодой подставки; арх – архегоний; ан – антеридий.

Рис. 12. Замшелый лес.
Гаметофит у всех Листостебельных мхов обычно радиально, реже двусторонне облиственный (рис. 13). От основания, а часто и вдоль стебля и от нижней части листьев отходят нитевидные, ветвящиеся, многоклеточные ризоиды. Листья сидячие, цельные, с жилкой или без нее. Стебель Бриевых симподиально или моноподиально разветвленный со сложным анатомическим строением, часто с дифференциацией на ткани, с центральным пучком или без него. На стебле образуются антеридии и архегонии.
Спорогон состоит из стопы, ножки (иногда ножка редуцирована) и коробочки. Спорообразующая ткань дает начало только спорам. Споры высвобождаются из коробочки, открывающейся в результате сбрасывания крышечки.


Рис. 13. Внешнее строение кукушкина льна.
Подкласс Сфагновые мхи – Sphagnidae
В этом подклассе всего 1 род сфагнум (Sphagnum), насчитывающий 300 видов растении бледно-зеленого цвета, за что их называют белыми мхами. Все представители класса – влаголюбивые растения, обладающие специальными приспособлениями для накопления и сохранения влаги. Стебель – прямостоячий, с густооблиственными боковыми ветвями, на верхушке ветви скучены в плотную головку, ниже собраны в пучки (рис. 14).


Рис. 14. Внешнее строение сфагнума.
Однослойные листья образованы из чередующихся клеток двух типов: узких живых фотосинтезирующих и широких мертвых водоносных (гиалиновых) (рис. 15). Из гиалиновых клеток состоит и прозрачная кожица, покрывающая стебельки и веточки (способность сфагнума впитывать воду в 20-40 раз больше). Ризоидов у взрослого растений нет, побег неограниченно долго нарастает верхней частью, постепенно отмирая снизу. Антеридии и архегонии образуются на особых ветвях, чаще ярко окрашенных. При созревании коробочка ссыхается, давление воздуха внутри нее повышается и крышечка резко, с хлопком отбрасывается. При этом из коробочки вылетает облачко спор на площадь до 10 см. Сфагнум формирует самые мощные торфяные залежи верховых болот.

Рис. 15. Клеточное строение сфагнума.
Представители Моховидных растут почти повсеместно. Известны так называемые моховые пустыни и тундры (рис. 16). До последнего времени совершенно не известны морские Моховидные, и только немногие виды найдены в зоне брызг на океанических побережьях. В то же время некоторые Моховидные (из родов бриум, меезия, дрепанокладус) могут произрастать на дне глубоководных (до 40 м) пресноводных водоемов. Гораздо больше болотных мхов. Обильно представлены мхи в среднеувлажненных и влажных лесах разных зональных типов, где они селятся на почве, камнях, валежнике, гнилой древесине, густо обрастают стволы и ветви деревьев.


Рис. 16. Распространение мхов.
В основном Моховидные не поедаются животными (могут быть побочным продуктом питания). Некоторые (сфагнум) обладают антибиотическими свойствами. Также используют торф – прессованные плиты для строительства, в сельском хозяйстве используют как топливо. Мхи аккумулируют многие вещества, в том числе радиоактивные. Моховидные играют важную роль в регулировании водного баланса континентов.
9








Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!

