
Россия, Кабардино-Балкария.


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Была в сети 28.03.2017 13:46

Отарова Елена Хазреталиевна
учитель
54 года
Местоположение
Специализация
Ткани высших растений.
Категория:
Биология
18.04.2016 00:42

Просмотр содержимого документа
«ткани высших растений.»
Занятие 4. Общая характеристика высших (Листостебельных (Наземных)) растений.
В царстве растений выделяют две большие группы: низшие и высшие растения. К низшим относят первично водные организмы – водоросли. У данной группы, как вам известно, тело представлено либо одной клеткой (хлорелла, эвглена), либо цепочкой клеток (нитчатые водоросли), либо слоевищем или талломом (рис. 1).
Под названием Высшие растения объединяются представители подцарства Листостебельные (Наземные) растения, сформировавшиеся в наземной среде и состоящие из побега (стебель, листья, почки) и корня. Такое листостебельное растение называется кормофит. Высшими эти растения называются потому, что в состав органов входят дифференцированные ткани. Существенной чертой Высших растений являются также многоклеточные гаметангии, причем, в большинстве отделов имеются специализированные женские гаметангии – архегонии.
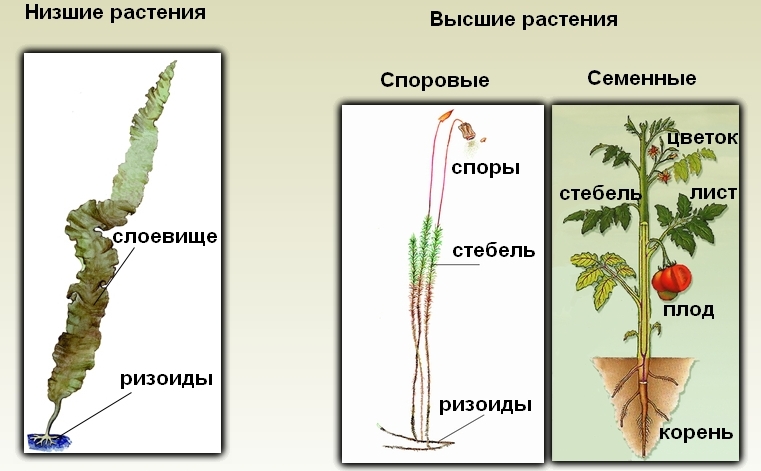
Рис. 1. Особенности строения низших и высших растений.
Первые наземные растения появились в силурийском периоде палеозойской эры (рис. 2). Большинство ботаников связывает их происхождение с вымершими формами зеленых водорослей. Предполагается, что толчком к выходу растений на сушу послужили: достаточное накопление в атмосфере свободного кислорода, усиление конкуренции в морях, появление озонового слоя, предохраняющего наземные растения от губительных ультрафиолетовых лучей. Первые наземные растения были травянистыми, но вскоре появились древовидные формы. Появление древовидных форм связано с соревнованием за верхние ярусы, где больше света.
Выход на сушу дал растениям ряд важных преимуществ – более доступные солнечный свет, кислород и углекислый газ, в результате чего в несколько раз ускорился фотосинтез. Но переход из водной среды на сушу был связан с многочисленными трудностями.
В водной среде растение не испытывает недостатка воды, которую оно может поглощать всей своей поверхностью. Наземные растения для защиты от высыхания вырабатывают защитную ткань – кожицу (эпидермис), которая выделяет на поверхность растения воскоподобную плёнку – кутикулу.
Поскольку кутикула сильно затрудняет жизненно важный газообмен между растением и окружающей средой, в кожице появляются специальные органы – устьица, состоящие из двух клеток, способных размыкаться, образуя пору для газообмена, и замыкаться на время, когда газообмен не нужен.

Рис. 2. Выход растений на сушу.
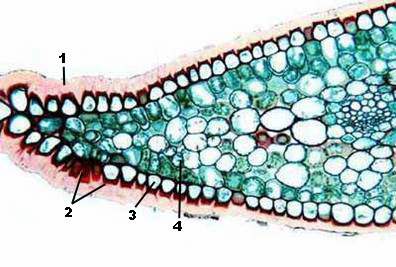
Развитие системы добычи воды из почвы привело к появлению корней, способных проникать на большую глубину.
Усовершенствование доставки воды из почвы в надземные части растения и синтезированных питательных веществ из надземных частей в подземные привело к появлению тканей сосудистой системы.
Для водных растений опорой служит вода. В воздушной среде с меньшей плотностью возникает потребность образования механических тканей для поддержания тела растения.
Совершенствуя систему улавливания света, сосудистые растения стали ветвиться. Сначала надземные части ветвились вильчато, затем у них стал различим основной стебель, с отходящими от него боковыми ответвлениями. При уплощении систем боковых ответвлений возникли пластинчатые органы – листья, позволяющие сильно увеличить поверхность фотосинтеза.
В водной среде не возникает сложностей при распространении спор, а в условиях суши необходимы приспособления для их рассеивания. В условиях суши большинство спор, рассеянных растением, гибнет, так и не попав в подходящие для прорастания условия, поэтому их нужно производить очень много. При этом споры защищены водонепроницаемой воскоподобной оболочкой.
Таким образом, развитие органов и тканей шло параллельно.
Тканью называют группу клеток, которые имеют одинаковое строение, происхождение и выполняют в организме сходные функции, а также межклеточное пространство (промежутки, образующиеся между клетками, называют). Не только клетки, но и межклетники входят в состав ткани.
Ткань называется простой, если все ее клетки одинаковы по форме и функциям (паренхима, склеренхима) (рис. 3). Сложные ткани (покровные, проводящие) состоят из клеток, неодинаковых по форме, внутреннему строению и функциям, но связанных общим происхождением.
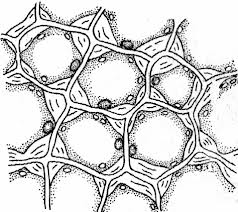
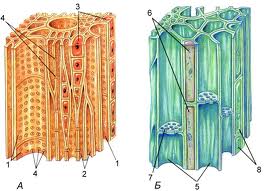
Рис. 3. Простая (склеренхима) и сложная (проводящая) ткани (слева направо)
По анатомо-физиологическому принципу выделяют образовательные, ассимиляционные, запасающие, воздухоносные, покровные, всасывающие, секреторные, механические и проводящие ткани.
Образовательные ткани (меристемы) являются эмбриональными. Благодаря долго сохраняющейся способности к делению (инициальные клетки, например, делятся в течение всей жизни) меристемы участвуют в образовании всех постоянных тканей, а также определяют рост и регенерацию растения.
Клетки меристем тонкостенные, многогранные, плотно сомкнутые, с густой цитоплазмой, тонкими оболочками, крупным ядром и очень мелкими вакуолями (рис. 4). Они способны делиться в разных направлениях. Если серединная пластинка перпендикулярна поверхности органа, такое деление называют антиклинальным. В результате образуется пластинчатая меристема (формирование листа). При заложении серединной пластинки параллельно поверхности органа возникает периклинальное деление, формирующее колончатую меристему (образование древесины, пробки). Деление с заложением перегородки касательно окружности определяют как тангенциальное. Клеточное деление при этом происходит во всех плоскостях, и в результате образуется меристема массы (образование спор, эндосперма).
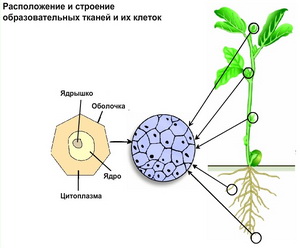
Рис 4. Строение и расположение образовательных тканей.
По происхождению меристемы бывают первичные и вторичные. Первичные образуются в результате дробления зиготы. Зародыш состоит только из нее. У взрослого растения она сохраняется только на кончике корней и верхушках побегов, что делает возможным их нарастание в длину. Вторичная (камбий и феллоген) развивается в онтогенезе от основной или покровной ткани либо первичной меристемы. Из вторичной развиваются все вторичные постоянные ткани, обеспечивая разрастание корня и стебля по диаметру (вторичный рост).
По местоположению меристемы можно разделить на апикальные, краевые, латеральные, интеркалярные и раневые.
Апикальные (верхушечные) располагаются на верхушке побегов и на кончике всех молодых корешков, обеспечивают рост растения в длину и называются конусом нарастания (рис. 5). У большинства растений здесь локализованы инициальные клетки, которые находятся в постоянном делении. В корне конус нарастания защищен корневым чехликом. В стебле – мелкими налегающими друг на друга листьями которые вместе с верхней частью стебля образуют почку. По происхождению верхушечные меристемы всегда первичны.
Топографически и онтогенетически с апикальной меристемой побега связано образование краевой (маргинальной) меристемы, формирующей пластинку листа.
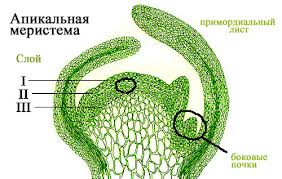
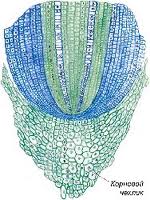
Рис 5. Апикальная меристема верхушки побега (слева) и кончика корня (справа).
Латеральные (боковые) способствуют росту растения в толщину и располагаются параллельно боковой поверхности того органа, в котором они находятся (рис. 6).
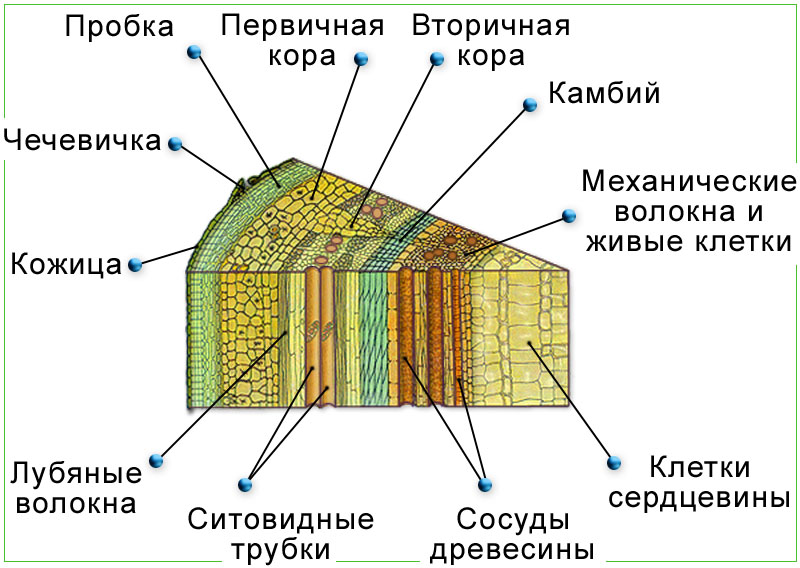
Рис. 6. Срез ствола дерева. Латеральная меристема (камбий).
Первичные латеральные меристемы (прокамбий, перицикл) возникают непосредственно под апексами и являются их производными. Вторичные меристемы (камбий и феллоген) образуются из тканей первичных меристем или из клеток постоянных тканей в процессе упрощения их структуры и приобретения свойств меристемы.
Интеркалярные (вставочные) располагаются обычно у основания междоузлий и обеспечивают рост растения в длину (рис. 7). Они имеют временный характер и превращаются в постоянные ткани.
Раневые (травматические) возникают в любой части растения при поранениях. Клетки постоянных тканей, окружающие повреждение, дедиффренцируются, приобретают способность к делению и образуют раневую ткань каллус (рис. 8). Клетки каллуса постепенно превращаются в клетки постоянной ткани (раневой пробки).
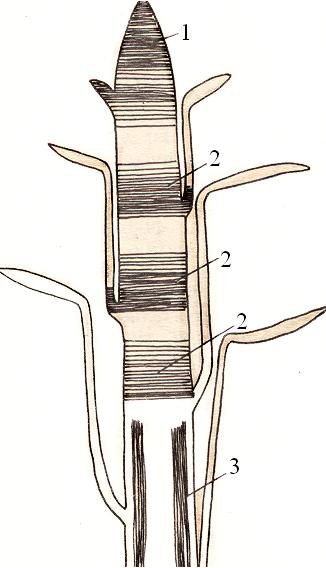

Рис. 7. Интеркалярная меристема (2). Рис. 8. Образование каллуса
Покровные ткани защищают внутренние ткани растений от прямого влияния факторов внешней среды, регулируют испарение и газообмен. В зависимости от происхождения различают три группы покровных тканей – эпидермис, перидерму и корку.
Эпидермис является сложной первичной покровной тканью и располагается на поверхности листьев и молодых стеблей. Основные клетки эпидермиса относительно неспециализированные и слагают основную массу ткани (рис. 9). Это живые, бесцветные, плотно прилегающие друг к другу клетки. Боковые стенки основных клеток часто бывают извилистыми, что повышает прочность их сцепления. Наружные стенки эпидермальных клеток наиболее утолщены и покрыты кутикулой. Поверх кутикулы обычно откладывается воск, придающий поверхности органа сизоватый оттенок. У некоторых растений (например, у хвойных) оболочки основных клеток одревесневают, а у хвощей – окремневают.
А Б
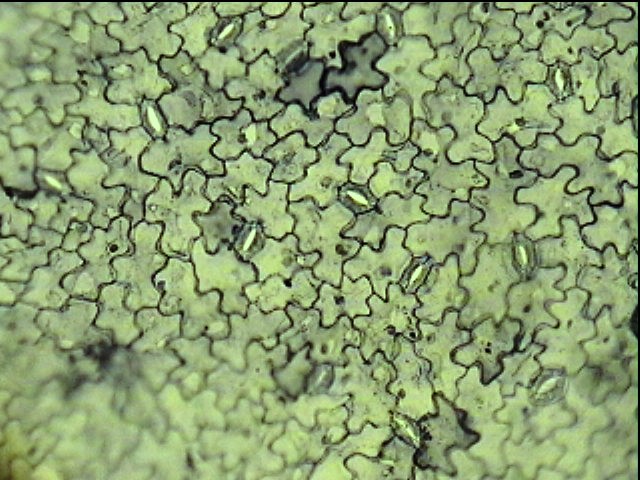
Рис. 9. Клетки эпидермиса листа. А – срез листа (1 – кутикула, 2 – утолщенные клеточные стенки, 3 – прозрачные клетки эпидермы, 4 – клетки мякоти листа); Б – поверхность листа.
В эпидерме листьев и зеленых стеблей имеются устьица, которые регулируют транспирацию и газообмен растения. Они представляют собой отверстия, ограниченные двумя замыкающими клетками. Рядом часто находятся побочные (обкладочные) клетки, отличающиеся от основных клеток эпидермиса размерами и формой и участвующие в движении устьиц. Замыкающие и побочные клетки составляют устьичный аппарат (рис. 10).
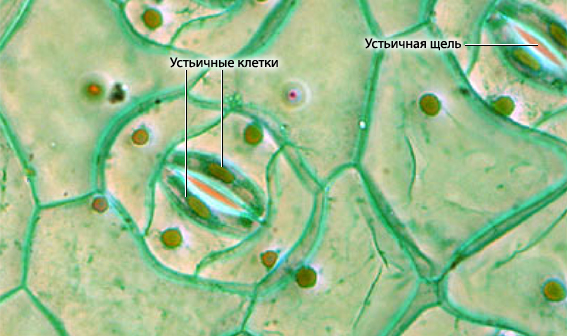
Рис. 10. Строение устьичного аппарата.
Замыкающие клетки всегда живые и содержат много хлоропластов, митохондрий и рибосом. Хлоропласты на свету активно фотосинтезируют, что приводит к накоплению органики и повышению осмотического давления. Это приводит к диффузии воды из окружающих клеток и к увеличению объема замыкающих клеток. При повышение тургора происходит растяжение более тонкой внешней стенки замыкающих клеток. Клетки искривляются и щель открывается. При потере воды клетки выпрямляются и щель замыкается. Устьица открываются на свету и к вечеру постепенно закрываются. В основном устьица располагаются на нижней поверхности листьев, что способствует меньшему испарению. У некоторых растений встречаются водные устьица или гидатоды которые секретируют воду.
Трихомы (волоски) представляют собой одно- и многоклеточные выросты эпидермиса (рис. 11). Если в образовании выроста задействованы ткани, лежащие под эпидермисом, такие выросты называют эмергенцами (шипы розы, волоски хмеля. Трихомы можно разделить на кроющие и железистые. Кроющие волоски – обычно мертвые образования, заполненные воздухом и покрывающие стебли и листья многих ксерофитов. Железистые волоски – живые структуры, выделяющие смолы, сахара, эфирные масла, слизи. К основным функциям трихомов относятся защита органов растений от перегрева, выведение токсичных солей из тканей листа, а также механическая и химическая защита от насекомых.
Рис. 11. Трихомы эпидермиса листа.
Эпиблема корня не является защитной тканью. Она выполняет функцию всасывания из почвы воды с растворенными веществами. Поэтому они имеют тонкую оболочку и обладают способностью образовывать корневые волоски (рис. 12). Для эпиблемы характерно отсутствие кутикулы и устьиц.

Рис. 12. Эпиблема корня.
Перидерма. Продолжительность жизни эпидермиса различна у разных видов и их органов. Например, на листьях и стеблях травянистых растений эпидермис сохраняется до конца их жизни. В стеблях и корнях, которые разрастаются в толщину путем вторичного роста, возникает вторичная покровная ткань – феллема (пробка). Вместе с феллогеном (пробковым камбием) и феллодермой она входит в состав перидермы, относимой в последнее время к особой анатомо-топографической зоне (рис. 13).
Феллоген представлен меристематической тканью, слоем прямоугольных клеток, уплощенных по радиусу органа. Феллоген внутрь откладывает клетки феллодермы, снаружи – пробки. Феллодерма представлена одним или несколькими слоями радиально расположенных паренхимных живых клеток, изнутри примыкающих к феллогену, и выполняет функцию его питания.
Молодые клетки феллемы (пробки), отложенные феллогеном, имеют тонкие оболочки. Затем возникают вторичные оболочки, содержащие ламеллы суберина и воска, вследствие чего их клеточная стенка опробковевает, теряет живое содержимое и заполняется воздухом. Пробка защищает растение от потери влаги, резких колебаний температуры, механических повреждений, микроорганизмов.
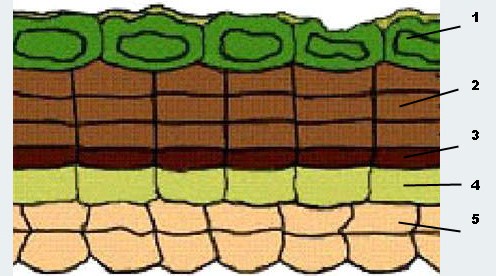
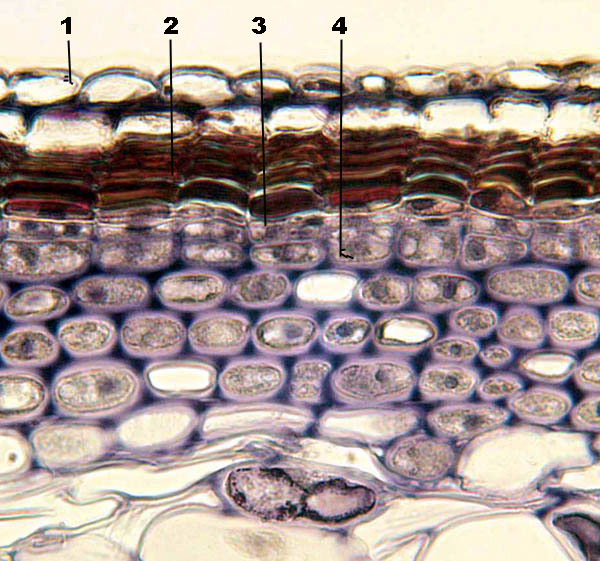
Рис. 13. Схема строения перидермы: 1 – слущивание клеток эпидермы с кутикулой, 2 – три слоя пробки, 3 – феллоген (пробковый камбий), 4 – феллодерма, 5 – первичная кора.
Лежащие под пробкой живые ткани испытывают потребность в газообмене. Поэтому в перидерме формируются чечевички – участки, через которые происходит газообмен (рис. 14).
Корка (ритидом) приходит на смену гладкой перидерме у некоторых древесных растений и состоит из чередующихся слоев перидермы и мертвой паренхимы, т. е. имеет сложный гистологический состав. Она предохраняет растение от механических повреждений, резких колебаний температуры, пожаров.
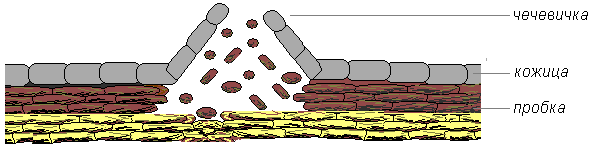
Рис. 14. Строение чечевички
Основные ткани (паренхима), состоит из живых, обычно тонкостенных клеток, которые составляют основу органов. В ней размещены механические, проводящие и другие постоянные ткани. Основная ткань выполняет ряд функций, в связи с чем различают ассимиляционную (хлоренхиму), запасающую, воздухоносную (аэренхиму) и водоносную паренхиму
Ассимиляционная ткань (хлоренхима) расположена под эпидермисом в листьях, неодревесневших стеблях, незрелых плодах, чашелистиках, т.е. в зеленых частях растения. Ее основная функция – фотосинтез. Клетки ассимиляционной ткани обычно паренхимные, тонкостенные, с большим количеством хлоропластов и межклетниками (рис. 15).
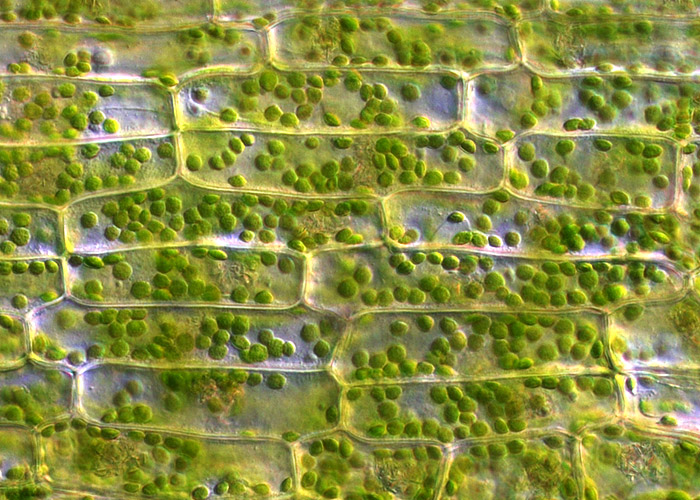
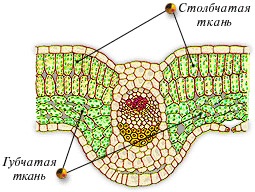
Рис. 15. Клетки хлоренхимы (слева), столбчатая и губчатая ассимиляционная ткань (справа).
Запасающие ткани представлены паренхимными тонкостенными клетками, в которых могут откладываться такие вещества, как крахмал, белки, сахара, жиры, вода (рис. 16). Данный тип тканей может быть локализован в различных органах растения (в семенах, корнях, клубнях, луковицах, корневищах, стеблях, листьях).
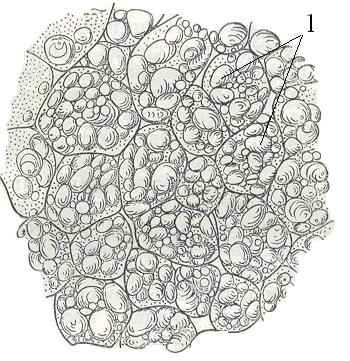
Рис. 16. Запасающая паренхима клубня картофеля. 1 – крахмальные зерна.
Воздухоносная ткань, или аэренхима, характеризуется наличием больших межклетников, осуществляющих газообмен и сообщающихся с внешней средой, как правило, посредством чечевичек и устьиц (рис. 17). Аэренхима хорошо развита у водных растений, а также видов, произрастающих на уплотненных и заболоченных почвах, где затруднено поглощение кислорода корнями.
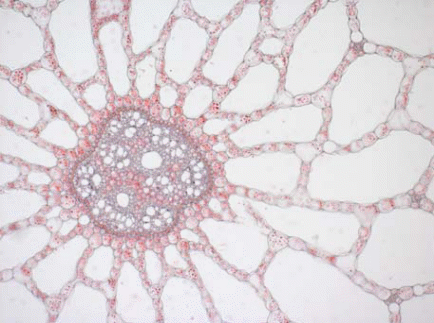
Рис. 17. Аэренхима в стебле рдеста.
Механические ткани обусловливают прочность растения. Стенки клеток, слагающих эти ткани, утолщены. Механические ткани чаще всего выполняют свое назначение в сочетании с остальными тканями растения, образуя их арматуру. К данному типу тканей относятся колленхима и склеренхима.
Колленхима – это механическая ткань, являющаяся первичной и служащая для укрепления молодых стеблей и листьев во время роста (рис. 18). Клетки колленхимы живые, с неравномерно утолщенными неодревесневшими стенками, вследствие чего они способны растягиваться при росте органа.
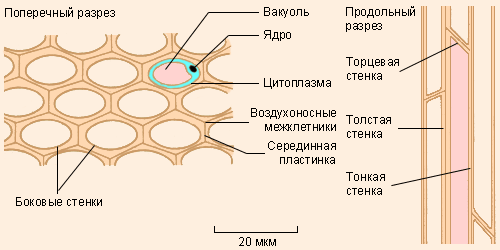
Рис. 18. Строение клеток колленхимы.
В зависимости от характера утолщения стенок различают уголковый (оболочек утолщается в углах, где сходятся 3-5 клеток), пластинчатый (тангенциальные стенки утолщаются сплошными параллельными слоями) и рыхлый (утолщение оболочек происходит на участках, примыкающих к межклетникам) типы колленхимы (рис. 19).
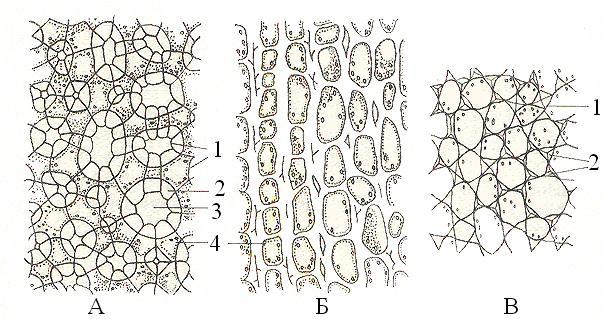
Рис. 19. Типы колленхимы (поперечный срез ): А – рыхлая; Б – пластинчатая; В – уголковая. 1 – первичная оболочка, 2 – утолщенная оболочка, 3 – межклетник, 4 – протопласт.
Склеренхима состоит из мертвых клеток с равномерно утолщенными и, как правило, лигнифицированными оболочками. Ее слагают два типа клеток: склеренхимные волокна и склереиды. Склеренхимные волокна образованы мертвыми клетками с острыми концами и толстыми оболочками, имеющими простые поры (рис. 20).
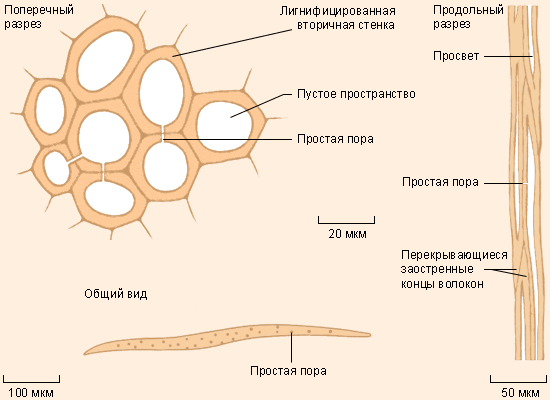
Рис. 20. Склеренхимные волокна.
Склереиды представляют собой мертвые клетки разнообразной формы с очень толстыми оболочками, пронизанными поровыми каналами. Клеточные стенки склереид одревесневают, в них откладываются известь, кремнезем, суберин, вследствие чего протопласт отмирает. Встречаются в плодах, листьях, стеблях, где располагаются поодиночке или группами (например, в мякоти плода груши). Склереиды, располагающиеся плотно, без межклетников, образуют косточки плодов сливы, вишни, абрикоса, скорлупу грецкого ореха.
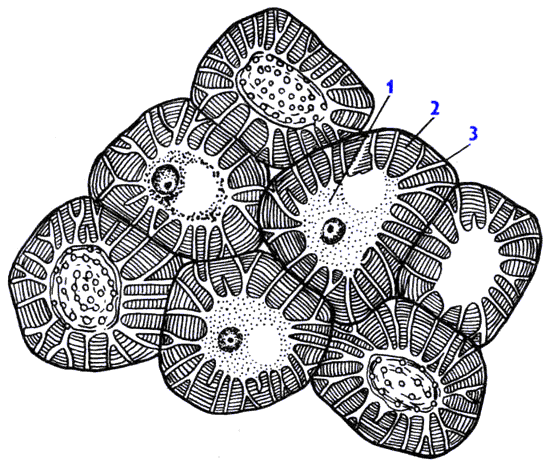
Рис. 21. Склереиды косточки созревающих плодов алычи с живым содержимым. 1 – цитоплазма, 2 – утолщенная клеточная оболочка, 3 – поровые канальцы.
Наряду с волокнами и склереидами, составляющими склеренхиму, в проводящей ткани высших растений имеются клетки, также специализирующиеся на выполнении опорной функции. Это древесинные (волокна либриформа) и лубяные волокна. Они отличаются от волокон типичной склеренхимы происхождением; поэтому рассматриваются как структурные элементы тех тканей, в которых образовались.








Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!

