
Россия, Республика Крым, Саки


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Была в сети 13.05.2025 11:51

Докаева Анастасия Сергеевна
учитель биологии
40 лет
Местоположение
Специализация
Докаева Анастасия Сергеевна
Категория:
Прочее
23.01.2019 16:17

Просмотр содержимого документа
«Докаева Анастасия Сергеевна»
ФИЗИОЛОГИЯ ЧЕЛОВЕКА TOM4-NS 1 1978
УД К 612.6.766.1
ВОЗРАСТНЫЕ ИЗМЕНЕНИЯ НЕКОТОРЫХ ПОКАЗАТЕЛЕЙ АЭРОБНОЙ ПРОИЗВОДИТЕЛЬНОСТИ У МАЛЬЧИКОВ 7—16 ЛЕТ 61-67 стр.
Л. А. Корниенко, Г. М. Маслова, В. Д. Сонькин, Л. Г. Евсеев
Научно-исследовательский институт физиологии детей и подростков АПН СССР,
Москва
Исследование развития тканевых окислительных систем в постна-тальном онтогенезе животных показало, что к периоду полового созревания происходит значительное увеличение аэробных возможностей скелетной мускулатуры, что обусловлено как нарастанием массы мышц, так и увеличением содержания в них важнейших окислительных ферментов [1, 2]. Исследование окислительных систем в тканях человека представляет известные трудности. В то же время известно, что в онтогенезе человека в пубертатный период возрастает работоспособность [3], резко увеличивается величина максимального потребления кислорода при физических нагрузках (МПК) [4—7]. Однако средние величины относительного МПК (рассчитанного на единицу веса тела), по-видимому, в течение длительного времени (от 6 до 25 лет) практически не меняются [3, 5]. Величины МПК в этом случае в большой степени зависят от тренированности и не характеризуют возрастные изменения аэробной производительности [3].
В ряде работ показано, что фактором, лимитирующим доставку кислорода к тканям, являются функциональные возможности сердца ![8, 9]. Поэтому многие авторы оценивают физическую работоспособность при работе определенной мощности по частоте сердечных сокращений [10— 14]. Широкое распространение получил тест PWC170 (Physical working capacity — физическая работоспособность) при пульсе 170 уд/мин, дающий представление о работоспособности при субмаксимальной нагрузке, близ верхнего порога аэробных возможностей мышц ,[12, 15]. В настоящее время этот тест рекомендован для применения в педиатрической практике [16], однако данных о возрастных изменениях iPWC в литературе мало.
В связи с этим мы предприняли попытку исследовать аэробную производительность у детей от 7 до 16 лет с использованием теста PWC170, полагая, что возрастные изменения функциональных возможностей сердца тесно связаны с развитием окислительных систем в мышечной ткани.
МЕТОДИКА
Исследовали PWC170 у 196 мальчиков в возрасте 7—16 лет, посещавших детский сад и школу. Нагрузку дозировали с 'помощью степ-теста и универсального велоэргометра конструкции Ю. П. Беликова в нашей модификации [17]. В обычной формуле для расчета работы на степ-тесте у мальчиков 7—8 лет мы использовали коэффициент 1, 2, вы-
61
как активность окислительных систем мышц до возраста 1'б лет, по-видимому, меняется незначительно. Это положение согласуется с данными ряда авторов, исследовавших кислородные режимы детей в онтогенезе [3,26].
ЗАКЛЮЧЕНИЕ
Полученные нами данные о возрастных изменениях физической работоспособности по тесту PWCm свидетельствуют об увеличении аэробной производительности организма в возрастном интервале от 7 до 16 лет. Наиболее значительные приросты отмечены в пубертатный период. Это увеличение связано в первую очередь с ростом сердца, что выражается в увеличении систолической работы и урежении пульса покоя. Величина PWC170, по-видимому, весьма информативна для оценки состояния ребенка в самых разнообразных ситуациях, в том числе для контроля за степенью адаптации организма к физической работе. Величина относительной PWC170 наряду с другими показателями может быть применена для более полной оценки физического развития детей и подростков.
Дополнительным показателем аэробных возможностей организма является .величина систолической работы (СР). Возрастные изменения относительной СР позволили предположить, что период наибольшего увеличения активности окислительных систем мышц приходится на возраст 7—9 лет. Предложенные данные позволяют считать этот период весьма благоприятным для развития аэробной производительности.
ЛИТЕРАТУРА
Маслова Г. М. Особенности тканевого дыхания, определяющие теплопродукцию
на различных этапах онтогенеза и при адаптации к холоду взрослых животных..
Канд. дис. М., 1971.
Корниенко И. А. и др. В сб.: Функциональные и адаптационные возможности детей
и подростков (тез конф. в Петрозаводске), т. 1. М., 1974, с, 30.
Колчинская А. 3. Кислородные режимы организма ребенка и подростка. Киев, 1973.
Robinson S. Arbeitsphysiologie, 1938, Bd 10, p. 251.
Astrand P. O. Experimental studies of physical working capacity. Copenhagen, 1952.
Фрейдберг И. М. Материалы V научн. конф. по возраста, морфол., фиэиол. и био
химии. М., 1961, с. 506.
Гуминский А. А. В кн.: Физическая культура и здоровье детей и подростков. М.,
1966.
Turino G. М. et al. J. Appl. Physiol., 1963, v. 18, p. 447.
Амосов Н. М., Бендет Я. А. Физическая активность и сердце. Киев, 1975.
Master А. М., Jafie Н. L. J. Mt. Sinai Host., 1949, v. 7, p. 629.
Lange Andersen K., Smith-Siversten A. In: Human adaptability and its methodology.
Tokyo, 1966, p. 183
Siostrand T. Acta med. scand., Suppl., 1947, v. 196, p. 687.
Wahlund H. Acta med. scand., 1948, v. 132, Suppl. 215.
Tornwall G. Acta physiol. scand., 1963, v. 58, Suppl. 201.
16. Карпман В. Л., Белоцерковский 3. Б., Гудков И. А. Исследование физической работоспособности у спортсменов. М., «Физкультура и спорт», 1974.
Иваницкая И. Н. Педиатрия, 1975, № 12, с. 62.
Евсеев Л. Г. В сб.: Проблемы возрастной физиологии. М., (1976, с. 81.
Roskamm Н. et al. J. Appl. Physiol., 1969, v. 27, p. 840.
Моно Г., Потье М. В кн.: Физиология труда. М., 11973.
Егоров П. А. В кн.: Тр. III научн. конф. по 'Возраста, морфол., физиол. и биохимии.
М., АПН, 1959, с. 237.
Кулаев Б. С., Бирюкович А. А. и др. Новые исследования по возрастной физиологии,
1976, № 1 (6).
Мякишев В. А. В сб.: Проблемы возрастной физиологии. М., 1976.
Holiday М. A. Pediatrics, 1971, v. 47, No. 1, 11, p. 169.
Калюжная Р. А. Физиология и и патология сердечно-сосудистой системы детей и
подростков. М., Медицина, 1973.
Пузик В. И., Харьков А. А. Возрастная морфология сердечно-сосудистой системы.
М., 1948.
Гуняди Б. К. и др. Новые исследования по возрастной физиологии; 1975, № 2.(5).
Абросимова Л. И., Карасик В. Е. Новые исследования по возрастной физиологии.
1077, №2 (9), с. 114.
Поступила в редакцию
29 июля 1976.
5 •
Костандов Э. А. Асимметрия зрительного восприятия и межполушарное взаимодействие // Физиология человека. – 1978. – Т.4. - №1. – С. 3-16
Научно-исследовательский институт судебной психиатрии им. В. П. Сербского, Москва
В настоящее время накапливается все больше данных, противоречащих мнению о полной специализации больших полушарий головного мозга человека, согласно которому определенная функция в них представлена унилатерально и может полноценно осуществляться соответствующим полушарием. Эта точка зрения в значительной мере опирается на клинико-морфологические данные, так как отмечаются несомненные корреляции между типом нарушения нервной или лсихической функции и поражением определенного полушария. Нередко встречающиеся случаи, частичного выпадения или восстановления функции сторонники теории специализации обычно объясняют неполным поражением соответствующих структур полушария или же атипичной морфофункцио--нальной организацией больших полушарий головного мозга (ярким выражением подобной атипичной организации, как известно, является леворукость).
Факты асимметрии зрительного и слухового восприятия, обнаруженные в многочисленных экспериментально-психологических работах последнего времени [1—8], также нередко рассматриваются как доказательство четкой полушарной специализации функций. Как известно, в такого рода экспериментах используются способы раздельного сенсорного стимулирования отдельных полушарий. Для слухового восприятия— это методика дихотичеокого .прослушивания, для зрительного — тахистоскопическая подача вербальных и невербальных стимулов на разные половины сетчатки. Принцип последнего методического приема основан на особенностях структурно-функциональной организации зрительной системы человека: объекты, находящиеся в левом поле зрения, раздражают правую половину сетчатки каждого глаза; нервные волокна из этих частей сетчатки обоих глаз приходят в зрительную кору правого полушария. Соответственно зрительная информация из правого поля зрения поступает в левое полушарие. Таким образом, эта методика дает возможность стимулировать зрительными импульсами непосредственно одно полушарие. Однако наблюдаемая в этих работах разница в порогах восприятия или в скрытых периодах реакции, очевидно, может быть объяснена не только задержкой или потерей информации при передаче через мозолистое тело из «неспециализированного» для данной функции полушария в соответствующее полушарие. Вполне законно предположение и о том, что некоторый дефицит функции при адресовании непосредственно в «неспециализированное» полушарие может быть обусловлен более медленной и менее совершенной обработкой инфор мации именно в этом полушарии. В качестве примера, подтверждающего такое объяснение, можно привести психологические исследования, проведенные [9—11] на людях с расщепленным мозгом.
Если при помощи тахистоскопа изолированно подавать отдельные слова, например, «ключ», «вилка», «кольцо», в правое полушарие, то человек с разобщенными полушариями не может- осознать и назвать их, но он в состоянии ощупью отобрать соответствующие предметы, которые эти слова обозначают. Следовательно, правое полушарие в известных пределах способно воспринимать и анализировать словесную информацию, однако для осознания ее необходимо, чтобы эта информация поступила в левое, так называемое доминантное полушарие, в котором находится «речедвигательный центра. Можно думать, что активация двигательной речевой области в процессе межполушарного взаимодействия является решающим звеном в структурно-функциональной мозговой системе, обеспечивающей осознание раздражителя.
Если семантический анализ и осознание внешнего сигнала осуществляются при непременном участии левого .полушария, то играет ли роль правое полушарие в обработке словесной информации? А если да, то в чем она заключается? Например, может ли полноценно осуществляться восприятие и осознание словесной информации при нарушении деятельности правого полушария? Выяснение этих вопросов, как нам кажется, имеет первостепенное значение для решения главной и принципиальной проблемы, а именно: существует ли полушарная специализация для определенной мозговой функции в целом или же для отдельных ее стадий, начиная от анализа физических свойств стимула, вербального и невербального, до корковой организации произвольных, сознательных реакций на него. В нашей лаборатории была проведена серия экспериментов, при помощи которых мы пытались ответить на вышепрставлен-ные вопросы.
Эксперимент 1 *. Для того 'чтобы свести к минимуму влияние на восприятие вербальных стимулов таких функциональных факторов, как приобретенный навык чтения слева направо, установка внимания и «деградация» следов зрительных запечатлений, и лучше выявить роль определенного полушария в осуществлении этой функции, мы использовали в качестве стимулов отдельные буквы, предъявляемые дихоптически с минимальной, околопороговой интенсивностью.
Исследование было проведено на 15 практически здоровых людях в возрасте от 19 до 45 лет и 14 больных с вялотекущим шизофреническим процессом без продуктивной психотической симптоматики.
Раздражители — изображения отдельных 'букв русского алфавита — предъявляются бинокулярно дихоптически, т. е. одновременно (разные буквы) в правом и левом полях зрения. Экран находится на расстоянии 80 см от глаз исследуемого. Размер каждой буквы равняется 17X15 мм; освещенность вблизи экрана 0,1 лк ±20%; послесвечение 1 мс; фиксационная точка (световое пятно) находится в центре экрана. Стимулы появляются на экране на 5 см вправо и влев/э от фиксационной точки (угол зрения — 3°36")- Перед каждым опытом дается инструкция: «Внимательно смотрите только перед собой на светящуюся точку. Голову не поворачивайте. Справа и слева от этой точки время от времени будут появляться буквы. Старайтесь их опознать. Как только это Вам удастся сделать, нажмите правой рукой на кнопку». Правильность опознания буквы определяется по словесному отчету после каждой пробы. Время экспозиции, вначале 10 мс, с каждым предъявлением стимулов увеличивается по 10 мс, пока исследуемый правильно опознает одну из букв в правом или левом поле зрения. В одном опыте определяется опознание трех-четырех пар букв. Буквы в разных опытах меняются местами в правом и левом полях зрения.
У здоровых исследуемых пороги опознания отдельных букв при их дихоптическом предъявлении равняются 20—40 мс. Они достоверно чаще
1 Эксперименты 1 и 2 проводились совместно с Г. С. Шостакович и Т. Н. Рещиковой. (pQ,Q5) опознаются в правом поле зрения, т. е. при их «прямом» адресовании в левое полушарие (в 77 случаях буквы опознавались а правом поле зрения и в 46 — в левом). В ряде случаев буквы одновременно опознавались в обоих полях зрения.
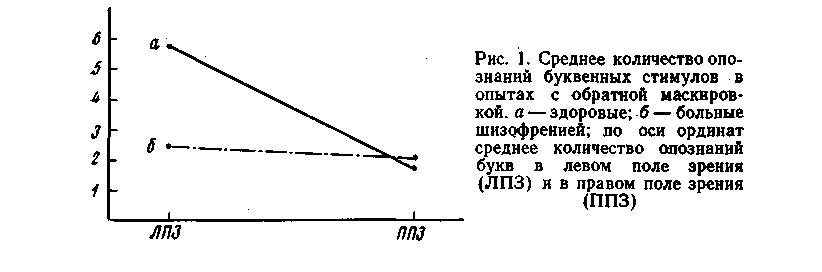
У больных шизофренией пороги опознания стимулов в два с лишним раза были выше, чем у здоровых лиц (50—80 мс). У них не проявляется преимущество поля зрения в опознании стимулов: из общего числа проб (169) отдельные буквы опознавались раньше в правом поле зрения в 39 случаях, в левом — в 32, в остальных пробах — одновременно в обоих полях зрения.
Полученные данные подтвердили уже известные факты о наличии асимметрии в зрительном восприятии вербальных стимулов, у здоровых людей [2-—5]. Как уже упоминалось, при наших условиях исследования функциональные факторы не могли играть существенной роли. Можно думать, что наблюдаемая асимметрия обусловлена особенностями реагирования левого полушария на «прямую» и «опосредованную» через правое полушарие зрительную стимуляцию. Как известно, информация при экспозиции стимула в правом поле зрения поступает непосредственно в левое полушарие, а из левого поля зрения — в правое полушарие. Весьма вероятно, что корковые структуры речевой зоны левого полушария легче возбуждаются при прямой стимуляции вербальными стимулами, чем при «опосредованной», так как в последнем случае происходит задержка или потеря информации при ее передаче через мозолистое тело из правого полушария в левое.
У больных шизофренией не отмечается асимметрии в опознании вербальных стимулов. Это нивелирование у них происходит на фоне значительного повышения порогов опдзнания стимулов, предъявляемых в правом и левом полях зрения. Мы не можем представить в настоящее время "удовлетворительного объяснения или гипртезы о нервных механизмах отмеченного факта — отсутствия у больных шизофренией асимметрии в опознании вербальных стимулов.
Эксперимент 2. При предъявлении раздражителя на .короткое время (до нескольких десятков миллисекунд) переработка информации не ограничивается периодом его экспозиции. Нервные следы стимула сохраняются и после окончания его действия, что, по-видимому, обеспечивает «считывание» информации в течение' времени, необходимого для осознания стимула [12]. Это видно, из того, что сильный маскирующий («стирающий») раздражитель, примененный спустя некоторое время после окончания экспозиции короткого тестового стимула, может помешать восприятию. Период времени с момента начала экспозиции короткого тестового стимула до включения маскирующего раздражителя, когда последний уже не может помешать осознанию тестового стимула, принято обозначать термином «время восприятия». Это время является показателем скорости поступления зрительной информации в кору больших полушарий, быстроты ее обработки-и сличения с образом, .хранящимся в памяти, в результате чего становится возможным осознание раздражителя. В феномене обратной маскировки участвуют как периферические, так и центральные, в частности корковые, нервные механизмы [13—15]. Таким образом, с помощью методики обратной маскировки измеряется время восприятия отдельных букв.
Тестовые стимулы и способ их предъявления, как в эксперименте 1. Исследуемые — те же лица, что в эксперименте 1.
Предварительно определяется порог эталонного опознания трех-четырех отдельных букв, в правом и левом полях зрения. Для этого процедура определения порога опознания одной и той же буквы повторяется многократно каждый раз, начиная с небольших экспозиций, до тех пор пока не устанавливается стабильное пороговое время для данной буквы (10—20 мс). После этого исследуется обратная маскировка. Ис-
пытуемый получает инструкцию: «Не обращайте внимания на возможные вспышки света. Ваша задача остается прежней — суметь прочесть буквы, которые появляются на экране справа и слева, и сообщить об этом нажатием кнопки».. В каждой пробе .для экспозиции стимулов в правое и левое поля зрения подбираются буквы с одинаковым эталонным порогом, и исследуемый не знает, какая буква предъявляется в данной пробе. Вслед за дихоптической экспозицией букв, порог которых был измерен в текущем опыте, через определенный промежуток времени включается маскирующий («стирающий») стимул — однократная вспышка лампы фотостимулятора «Нихон-Коден» интенсивностью 0,27 Дж, длительностью 100 мкв. Лампа фотостимулятора расположена в поле
ясного видения исследуемого. Маскирующий стимул включается через 20 мс с момента окончания экспозиции буквы. Затем пауза между тестовым и маскирующим стимулами с каждой пробой увеличивается по 10 мс, до тех пор пока исследуемый опознает букву в одном из полей зрения. В одном опыте таким способом измеряется время восприятия двух-трех букв.
В условиях обратной маскировки здоровые исследуемые достоверно чаще опознают буквенные стимулы в левом поле зрения: в 85 пробах буквы сначала опознавались в левом поле зрения и только в 28 пробах— в правом (р
У здоровых взрослых людей, как показывают наши данные, при ди-хоптическом способе предъявления отдельные буквы в условиях обратной маскировки достоверно чаще распознаются в левом поле зрения, чем в правом. Время восприятия, т. е. время, необходимое для передачи зрительной вербальной информации в кору и ее дальнейшей переработки, в результате чего происходит осознание этой информации, короче, когда стимул расположен в левом поле зрения (по сравнению -с правым полем зрения). Этот факт может быть объяснен с точки зрения концепции о «сотрудничестве» двух полушарий. 'Умозрительно можно представить следующую схему обработки зрительной информации при экспозиции буквы в левом поле зрения. Зрительные имлульсы поступают сперва в правое полушарие, где, по-видимому, осуществляется зрительно-пространственный анализ стимула, а затем информация передается в
левое полушарие. В последнем происходит уже высший анализ и синтез вербальных признаков стимула и его осознание. Вероятно, это более коррткий путь последовательных этапов переработки зрительной информации, чем при адресовании ее в левое «речевое» полушарие. В последнем случае информация уже опосредованно поступает в правое полушарие, там осуществляется зрительно-пространственный анализ элементов буквы, а затем возвращается в переработанном виде в левое полушарие, где уже окончательно -происходит ее семантический анализ. Поэтому и время, необходимое на обработку и осознание вербальной информации, несколько больше при «непосредственной» стимуляции левого полушария по сравнению с подачей стимула «прямо» в правое полушарие.
Другое возможное объяснение асимметрии времени восприятия, не исключающее первое, опирается на электрофизиологические данные, полученные в нашей лаборатории. Анализ временных параметров корковой вызванной электрической активности дает основание считать, что правое полушарие быстрее, чем левое, обрабатывает информацию. С этой точки зрения, надо думать, у больных шизофренией теряется отмеченное преимущество правого полушария в быстроте обработки зрительной информации.
Эксперимент 3. Исследовали 10 здоровых человек, правшей, с нормальным зрением, в возрасте от 19 до 45 лет.
Попытки выявить полушарную асимметрию по форме или временным и амплитудным параметрам вызванных корковых потенциалов на вербальные и невербальные раздражители не дали однозначных результатов [16—22]. Возможно, это обусловлено тем, что не во всех подобных работах применялись методы сенсорной стимуляции, принятые в экспериментальной психологии при исследованиях асимметрии функций полушарий головного мозга. В наших совместно с О. А. Генкиной экспериментах использовалась тахистоскопическая методика, позволяющая подавать зрительную информацию изолированно в правое или левое полушарие путем кратковременного предъявления раздр-ажителя попеременно в левом или правом поле зрения-.
Исследуемый во время опыта полулежит с открытыми глазами в специальном кресле в звукозаглушенной кабине. Зрительные стимулы — изображения отдельных букв русского алфавита или «бессмысленных» фигур. Буквенные стимулы мы называем «вербальными», поскольку они являются элементом речи. Мы отдаем себе отчет в некоторой условности этого обозначения, так как буква не несет семантической информации. «Бессмысленными» фигурами мы, в согласии с литературными данными [3], называем изображения фигур сложной, неправильной, случайной конфигурации, которую трудно вербализовать. .После каждого опыта исследуемому показывают весь набор изображений бессмысленных фигур, и он должен опознать те из них, которые предъявлялись в данном опыте. Вербальные и невербальные раздражители .предъявляются ритмически — один раз в 3 с, вразбивку, в случайном порядке в разные половины поля зрения, на 31 см вправо или влево от фиксационной точки в центре экрана (угол зрения равен 20°). Экран находится на расстоянии 80 см от глаз исследуемого. Время экспозиции стимулов 10 мс. Короткая экспозиция стимулов была выбрана по двум причинам: во-первых, при такой длительности исследуемый не .успевает перевести взор от фиксационной точки в сторону стимула; во-вторых, имеются данные о том, что полушарная асимметрия лучше выявляется при использовании коротких зрительных стимулов [1, 23].
Размер каждого стимула — буквы или фигуры— 17x15 мм. Площадь высвечиваемых на темном фоне электролюминесцентного экрана фигур и букв одинаковая. Цвет свечения белый, послесвечение 1 мс. Освещенность вблизи экрана 0,1 лк±20%. Все стимулы легко опознаются иссле-
7
дуемым, о чем мы судим по его словесному отчету после каждой пробы. Движения глаз фиксируются на электроэнцефалографе. Опыт начинается после 5-минутной адаптации к темноте. Предварительно исследуемому дается инструкция? «Внимательно смотрите только перед собой на светящуюся точку; голову не поворачивайте; справа и слева от точки время от, времени будут появляться буквы или изображения фигур, старайтесь их опознать и запомнить, сколько раз они появятся».
Электрическая активность регистрируется, монополярно. Активные электроды помещаются на левой и правой затылочных областях черепа (на 2,5—3 см выше и на 3 см левее и правее затылочного бугра), индифферентный электрод — на мочке левого уха. Потенциалы через усилители электроэнцефалографа «Нихон Коден» ME-132 ('постоянная времени 0,3 с) отводятся на магнитное регистрирующее устройство SDR-813 и в последующем усредняются при помощи специализированной ЭВМ APT-1000 раздельно в зависимости от подачи стимула в правую или левую половины поля зрения. В каждой пробе усредняются 50 биоэлектрических реакций на стимулы, экспонируемые в левой половине поля зрения, и 50 реакций на стимулы — в правой. Усредненные вызванные потенциалы (УВП) фотографируются с экрана осциллоскопа APT-1000. В данной работе анализируется латентный период (ЛП) и амплитуда пика поздних компонентов УВП — отрицательного N200 с ЛП около 200 мс и положительного Р 300-—с ЛП около 300 мс. Результаты всех измерений обрабатываются статистически. Всего взято в обработку 136 УВП — на буквенные стимулы и 111 — на фигуры.
Различие в реакциях левого и правого полушарий на «прямое» раздражение одними и теми же буквенными стимулами (правое поле зрения — левое полушарие; левое поле зрения — правое полушарие) выявляется при анализе ЛП поздних компонентов.УВП (рис. 2). Разница в ЛП особенно четко видна при сопоставлении компонентов Р 300 (при статистической обработке методом сопряженных пар — р
Существенная разница в реакциях на вербальные и невербальные раздражители наблюдается в пробах, где они предъявляются в левом поле зрения («прямое» — правое полушарие). В этих случаях с правой затылочной области на буквенный стимул регистрируется компонент Р 300, скрытый период которого в среднем составляет 276±2,7 мс, а на изображение фигуры — 267±3,0 мс (р0,2).
Межполушарная асимметрия выявляется также в пробах, в которых зрительное раздражение адресуется в одно полушарие, а вызванный потенциал отводится с противоположного полушария. Как можно видеть на рис. 3, при опосредованном — через левое полушарие — зрительном раздражении (буквенный стимул) поздний положительный компонент Р 300 в правом полушарии развивается с большим скрытым периодом, чем аналогичный потенциал в симметричной левой затылочной области, когда раздражение поступает сначала в правое полушарие (290±3,3 и 280±2,9 мс соответственно; р
Итак, независимо от характера зрительного раздражителя поздний положительный вызванный потенциал в левом полушарии при «прямом» раздражении развивается с достоверно большим скрытым периодом, чем в правом полушарии. В правом полушарии этот ответ на изображение фигуры развивается раньше, чем на буквенный раздражитель. При опосредованном через правое полушарие раздражении поздний положительный потенциал в левой затылочной области развивается с меньшим скрытым периодом, чем в правой затылочной области, когда сенсорная стимуляция «прямо» подается в левое полушарие.
На рис. 4 представлена величина поздних вызванных электрических ответов обоих полушарий в пробах, где каждое из них раздражается в случайном порядке путем предъявления стимула в соответствующем
I'm:, о
контралатеральном поле зрения. На буквенный стимул в левом полушарии амплитуда компонента Р 300 достоверно больше, чем в правом (р
Величина компонента Р 300 в левом полушарии на стимул в правом поле зрения («прямое»'—левое полушарие) достоверно больше (р
Итак, у здоровых исследуемых поздний положительный компонент Р 300 УВП существенно больше по величине в левом полушарии, чем в правом, независимо от характера раздражителя, предъявляемого в
контралатеральном поле зрения. Только в левом полушарии наблюдается достоверная разница в величине волны Р 300 между «прямым» и «опосредованным» способами предъявления стимулов.
Анализ скрытых периодов вызванных потенциалов правой и левой корковых затылочных областей на стимулы, подаваемые в разные половины поля зрения, дает возможность судить о некоторых временных параметрах межполушарного взаимодействия у человека. При «опосредованной» сенсорной стимуляции (раздражитель подается в ипсилатеральном поле зрения, т. е. прямо адресуется в одно полушарие, а потенциал отводится с другого) вызванный потенциал с левой затылочной области отводится с более коротким скрытым периодом, чем с правой симметричной области, когда последняя раздражается таким же способом. Эта разница больше выражена в пробах с невербальными стимулами. На основании этих данных можно было бы считать, что левое полушарие реагирует быстрее, чем правое. Однако пробы с «прямой» стимуляцией отдельного полушария, когда стимул подается в контралатеральном поле зрения, показывают, что скрытый период коркового ответа в левом полушарии явно больше, чем в правом. Таким образом, если судить по электрическому вызванному ответу, левое полушарие реагирует медленнее, чем правое. Этот факт делает вероятным другое предположение: более короткий скрытый период коркового ответа в левом полушарии при стимуляции в ипсилатеральном поле зрения (опосредованная стимуляция) обусловлен тем, что нервные импульсы из «прямого» правого полушария поступают в левое быстрее, чем из левого полушария (в пробах, где оно «прямое») в правое. По всей вероят-
ности, это происходит потому, что в правом полушарии быстрее, чем в левом, организуется вызванный корковый ответ,.а следовательно, передача в противоположное полушарие нервных импульсов, участвующих в генезе поздних волн, отсюда осуществляется быстрее, чем из левого полушария в правое.
Очевидно, в процессе межполушарного взаимодействия осуществляется передача не только специфической зрительной информации, но и результатов ее корковой переработки. Об этом говорят следующие факты. Скрытый период позднего вызванного ответа Р 300 в правом полушарии на «прямую» стимуляцию значительно короче на невербальные стимулы, чем на буквенные. Такая же разница в этой пробе наблюдается и в левом, «опосредованном» полушарии. При «прямой» стимуляции левого полушария подобной разницы во временных параметрах УВП на невербальные и вербальные стимулы не наблюдается ни в этом, ни в противоположном полушарии.
Таким образом, наши данные подтверждают точку зрения, согласно которой через мозолистое тело из одного полушария в другое передается не только (или даже не столько) специфическая зрительная информация, но и сложный нейронный код уже образованной временной связи или ассоциации, т. е. обучения и памяти [24]. На это указывают морфо-физиологические факты. Было показано, что нижневисочная кора, которая играет .важную роль в зрительном обучении, связана через мозолистое тело и переднюю комиссуру не только с престриарной зоной, но и с нижневисочной областью противоположного полущария [25].
Итак, электрофизиологические данные говорят о правомерности сделанного-нами в разделе «Эксперимент 2» предположения о том, что в процессе межполушарного взаимодействия из правого полушария в левое передаются результаты зрительно-пространственного анализа вербального стимула. Подобный анализ и передача его результатов в левое «речевое* полушарие, по всей вероятности, 'являются необходимым этапом в сложном процессе восприятия вербальных стимулов. При этом Эффективность межполушарного взаимодействия, по-видимому, обеспечивается тем, что в правом полушарии обработка информации, особенно невербальной, осуществляется быстрее, чем в левом.
Полушарная асимметрия, выявляемая при анализе амплитуды вызванных потенциалов на вербальные и невербальные стимулы, говорит о преимущественной и только относительной функциональной специализации левого или правого полушария-. Существенной внутриполушарной разницы в величине вызванного ответа на вербальные и невербальные стимулы не отмечается ни в левом, ни в правом полушариях. Разница в амплитуде УВП на оба вида раздражителей в основном межполу-шарная.
Как известно, происхождение позднего положительного компонента Р 300 у человека связано с ассоциативными процессами в неокортексе [26]. У бодрствующего нормального человека эти процессы осуществляются при участии второй сигнальной системы. Очевидно, этим следует объяснить наблюдаемый в наших опытах факт большей выраженности волны Р 300 в левом «доминантном» полушарии, которое в значительно большей степени, чем правое полушарие, связано с речевой функцией. Только в левом полушарии выявляется разница в величине вызванного коркового ответа на «прямое» и «опосредованное» раздражения. Однако следует отметить, что эта разница отмечается как на, вербальные, так и яа невербальные стимулы.
С другой стороны, наблюдаемая в наших исследованиях полушарная асимметрия в вызванной электрической активности коры головного мозга в виде большей амплитуды позднего отрицательного колебания в правом полушарии на невербальные стимулы является непосредственным доказательством определенной функциональной «специализации» правого полушария для восприятия и переработки несловесной зрительной4 информации.
Эксперимент 4. Регистрация вызванной электрической активности в
эксперименте 3 производилась, по понятным причинам, в условиягх уни-
латерального предъявления зрительных стимулов. Поэтому, мы с
О. А. Генкиной считали необходимым — в целях сопоставления биоэлек
трических данных с показателями зрительного восприятия — исследовать-
пороги опознания вербальных стимулов при таком же способе стиму
ляции. . . .,
Исследование проведено на .практически здоровых людях (16 человек; 10 из них исследовались в эксперименте 3). Характер и способ-предъявления зрительных раздражителей (величина стимулов, освещенность, цвет, расстояние экрана от глаз исследуемого) те же, что и в эксперименте 3, за исключением того, что в качестве невербальных используются изображения геометрических фигур, например, треугольник, ромб, квадрат. Время экспозиции каждого стимула во всех опытах вначале заведомо небольшое (10 мс), так что исследуемый не может его опознать. Затем это .время с каждым предъявлением стимула увеличивается на 5—10 мс, до тех пор пока исследуемый его опознает и правильно называет вслух. Перед каждым опытом дается инструкция: «Внимательно смотрите .только перед собой на светящуюся точку; голову не поворачивайте; справа или слева от точки время от времени будут появляться буквы или изображения фигур. Старайтесь их опознать. Как только это Вам удастся сделать, произнесите вслух обозначение буквы или фигуры». В каждом опыте определяется порог опознания двух-трех букв или фигур в правом и столько же в левом полях зрения.
Порог опознания отдельных букв в левом поле зрения в среднем равняется 59±2,5 мс, в правом — 52±2,2 ис (pQ,Q5). Он почти в 3 раза чаще был ниже в пробах, где буквенный стимул предъявлялся в правом поле зрения.
При опознании невербальных стимулов (изображения геометрических фигур) не отмечается существенной разницы в зависимости от поля зрения (в левом поле зрения порог опознания в среднем равняется 38 + ±1,7 мс, в правом —36± 1,4 мс; р0,3).
Итак, при унилатеральном способе подачи зрительных стимулов определенные буквы опознаются значительно лучше в правом поле зрения, в то время как изображения геометрических фигур воспринимаются по существу одинаково в обеих полях зрения.
При унилатеральном способе предъявления зрительных сигналов роль таких функциональных факторов, как закрепленный в течение жизни навык чтения слева направо, установка внимания и «деградация» следа зрительных запечатлений, не играют существенной роли [5]. Необходимо принять во внимание фактор доминантности глаза, который особенно должен влиять на восприятие стимулов малой интенсивности, как это было в наших опытах. Однако если латерализация 'остроты зрения играла бы определенную роль, то это обстоятельство отразилось бы и на восприятии невербальных стимулов. Между тем асимметрии опознания геометрических фигур не наблюдается.
По всей вероятности, наблюдаемая в наших исследованиях асимметрия в опознании вербальных стимулов в основном определяется особенностями реагирования левого полушария. Как показал анализ вызванных потенциалов, проведенный в разделе «Эксперимент 4», левое полуг шарие значительно более чувствительно к «прямой» зрительной стимуляции, чем к «опосредованной», поступающей из левой половины поля зрения. Этой разницы нет в правом полушарии при аналогичных способах его раздражения.
Вышеприведенное объяснение нельзя считать полностью удовлетворительным, так как величина вызванных потенциалов на «прямое» раздражение левого полушария больше, чем на «непрямое», не только на буквенные стимулы, но и на невербальные раздражители. В то же время асимметрия порогов опознания наблюдается только на вербальные стимулы.
Эксперимент 5. У здоровых людей регистрация зрительных вызванных потенциалов выявила определенную межлолушарную разницу в процессе восприятия вербальных и невербальных раздражителей. С целью выяснения функционального значения для высшей нервной деятельности человека этой разницы О. А. Генкина исследовала межполу-шарные отношения у лиц с патологически низким интеллектом — у олигофренов в легкой и средней степени дебильности. Это исследование оправдано тем, что, согласно некоторым клинико-психологическим данным, у лиц с врожденной умственной отсталостью слабо выражена или же вовсе отсутствует латерализация функций [27]. Среди них относительно мало праворуких, не выражена доминантность правого глаза и т. д. Однако следует заметить, что лево- или праворукость не является достаточным показателем полушарной речи и тем более далеко не все леворукие обнаруживают расстройства речи или интеллекта и наоборот.
Имеются экспериментально-психологические данные о том, что у школьников с низким умственным развитием асимметрия восприятия вербальных стимулов больше выражена, чем у нормальных детей того же возраста, за счет худшего опознания раздражителей в левом поле зрения, т. е. определенного дефицита функций правого полушария [28]. Вместе с тем у умственно отсталых детей в отличие от здоровых не обнаружено асимметрии в вызванной электрической активности на невербальные зрительные раздражители [28, 30]. Таким образом, в литературе имеются довольно противоречивые и неопределенные сведения о связи полушарной асимметрии функций с уровнем интеллекта.
Исследование проведено на шести больных олигофренией в степени дебильности, с нормальным зрением, праворуких, в возрасте 18—20 лет. Методика исследования та же, что в эксперименте 3.
У лиц с умственной отсталостью, как и у нормальных исследуемых, скрытый период поздних компонентов вызванного потенциала на «прямое» раздражение буквенными стимулами в левом полушарии больше, чем в правом (рис. 2). Эта разница достоверна как по позднему отрицательному компоненту N 200, так и по положительному компоненту Р 300 (при статистической обработке методом сопряженных пар по обоим компонентам — р
Как описывалось выше, на невербальные стимулы у здоровых исследуемых, как и на буквенные, отмечается разница в скрытом периоде поздних волн при «прямой» стимуляции левого и правого полушарий (рис. 2). У умственно отсталых исследуемых достоверной разницы в скрытом периоде вызванных ответов на «прямое» раздражение невербальными стимулами не отмечается (р;0,1).
Если сравнить скрытые периоды вызванного ответа' Р 300 в правом полушарии на его «прямое» раздражение буквенными и невербальными стимулами (стимул в левом поле зрения), то у умственно отсталых исследуемых разницы не отмечается (р0,4). У здоровых людей вызванный ответ на невербальные раздражители в правом полушарии развивается достоверно раньше, чем в левом (см. эксперимент 3).
При опосредованной зрительной стимуляции, когда раздражитель адресуется в одно полушарие, а потенциал отводится с противоположного (рис. 3), у олигофренов в отличие от здоровых испытуемых не отмечается существенной разницы в скрытом периоде поздней положительной волны Р 300 как на вербальные стимулы, так и на изображения «бессмысленных» фигур (р0,3).
Сравнение скрытых периодов позднего отрицательного компонента .N 200 и позднего положительного компонента Р 300 у здоровых и олигофренов показывает, что у последних корковый ответ явно замедлен* Большее скрытое время вызванного ответа у умственно отсталых отмечается как при «прямой», так и «опосредованной» стимуляциях левого и правого полушарий, как на вербальные, так и невербальные зрительные раздражители. Во -всех случаях эта разница статистически высоко* достоверна (р
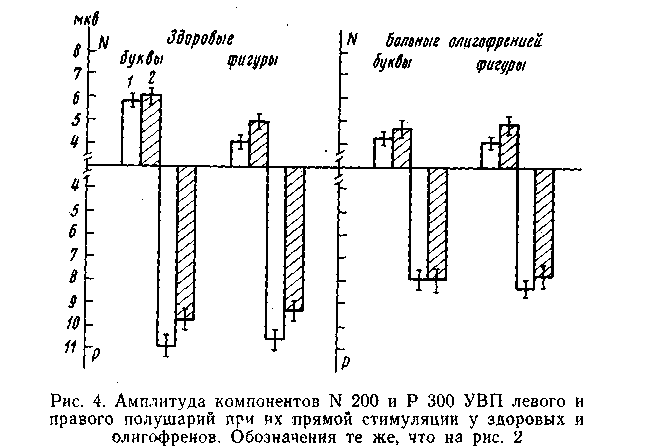
Амплитуда поздних вызванных ответов у олигофренов существенно меньше, чем у здоровых. На рис. 4 видно, что у умственно отсталых исследуемых амплитуда компонентов N 200 и Р 300 сравнительно невелика в обоих полушариях как на буквенные, так и на невербальные стимулы. Другое отличие от здоровых заключается в отсутствии у умственно отсталых исследуемых полушарной асимметрии в величине позднего компонента Р 300 вызванного потенциала как на буквенные, так и. на невербальные раздражители (рис. 4).
Как описывалось выше (эксперимент 3), у здоровых людей в левом полушарии наблюдается достоверная разница в величине волны Р 300 на «прямой» и «опосредованный» способы предъйвления стимулов — на раздражитель в правом поле зрения потенциал существенно больше, чем на тот же стимул в ипсилатеральнсм поле зрения. У олигофренов подобной разницы не наблюдается. Например, на буквенный стимул в правом поле зрения с «прямого» левого полушария регистрируется потенциал Р 300, амплитуда которого в среднем равняется 7,9±0,37 мкВ^ а на тот же стимул, экспонируемый в левом поле зрения, с этого же места отводится потенциал с амплитудой 7,6±0,37 мкВ, т. е. никакой разницы в величине Р 300 между «прямым» и «опосредованным» раздражениями левого полушария у олигофренов не отмечается в отличие-от здоровых.
Итак, у умственно отсталых лиц скрытый период поздних вызванных ответов (компоненты N 200 и Р 300) больше, а их амплитуда явно меньше, чем у здоровых исследуемых. У олигофренов вызванный ответ в правом полушарии на невербальную стимуляцию не развивается, как: у здоровых исследуемых, с более коротким скрытым периодом, чем на вербальную. У олигофренов не отмечается межтюлушарной разницы в амплитуде поздних волн вызванного потенциала- как на буквенные, так и на невербальные стимулы.
Полученные О. А. Генкиной данные показывают, что у олигофренов соотношения временных и амплитудных параметров поздних вызванных потенциалов между левым и правым полушариями существенно отличаются от здоровых исследуемых. Это прежде всего выражается в одинаковом скрытом периоде вызванных потенциалов в обоих полушариях на невербальные раздражители у олигофренов. У них же в правом полушарии отсутствует разница в скрытом периоде позднего положительного компонента Р 300 на вербальные и невербальные стимулы. Как было показано нами выше, у здоровых людей корковый вызванный ответ, особенно на невербальные раздражители, в правом полушарии организуется раньше, чем в левом. По нашему мнению, в этом заключается большой функциональный смысл: для успешного коркового анализа и синтеза собственно речевого сигнала необходим предварительный анализ И синтез его физических параметров, что и осуществляется преимущественно в правом полушарии за счет более быстрой обработки поступающей информации. У олигофренов отсутствует подобное опережение-обработки информации в правом полушарии, что, возможио, является одной из причин нарушений у них процессов переработки речевой информации. Можно предположить, что это один из вероятных нервных механизмов снижения мыслительной функции, которая, как известно, у человека осуществляется на базе речевой деятельности.
Другое отличие вызванной активности левого и правого полушарий, выявляемое в процессе восприятия речевых сигналов, связано с амплитудой позднего положительного компонента Р 300. Волна Р 300, отражающая корковые ассоциативные познавательные процессы, у здорового человека при прямой стимуляции лучше выражена в левом полушарии независимо от рода стимула (вербальные или невербальные). У умственно отсталых исследуемых не отмечается подобной межполу-шарной асимметрии в амплитуде волны Р 300. При этом величина ее существенно меньше, чем у нормальных лиц. Вышеотмеченный факт как нам кажется, является показателем патологических отклонений в деятельности левого «речевого» полушария. Можно думать, что у олигофренов внешний сигнал вызывает возбуждение меньшего по сравнению с нормой числа корковых элементов обоих полушарий (особенно левого), принимающих участие в обработке поступающей информации, что, несомненно, должно сказаться на быстроте анализа и синтеза этой информации и их качестве.
ЗАКЛЮЧЕНИЕ
Проведенные в нашей лаборатории исследования выявили определенную асимметрию зрительного восприятия вербальных и невербальных стимулов, которая отражалась на порогах опознания и в амплитудно-временных параметрах корковой вызванной активности. Вместе с тем целый ряд полученных нами фактов можно объяснить в свете концепции о взаимодополняющем «сотрудничестве» двух полушарий в процессе осуществления целостной функции восприятий. Так, например, оказалось, что время восприятия, т. е. время, необходимое на переработку и осознание вербальной информации, короче, если она «прямо» адресуется в правое полушарие. Очевидно, начальный этап переработки вербальной информации, а именно зрительно-пространственный анализ элементов буквы, осуществляется преимущественно в правом полушарии. Сделанное предположение опирается на следующий факт: скрытый период вызванного коркового ответа в правом полушарии короче, чем в левом, при их «прямой» стимуляции.
В правом полушарии быстрее, чем в левом, организуется вызванный корковый ответ, чем обеспечивается быстрая передача результатов корковой обработки информации в противоположное полушарие. По-видимому, этим достигается временная последовательность этапов корковой переработки вербальной информации, которая начинается с анализа физических признаков отдельных букв в правом полушарии, а затем продолжается в левом, где уже происходит семантический анализ. Так, например, различение букв только по физическим параметрам (предъявляется однг. и та же буква, но разной величины) быстрее осуществляется, если они находятся в левом поле зрения, т. е. информация «прямо» подается в правое полушарие. Если же исследуемый должен опознавать разные буквы не по физическим характеристикам, то четко выявляется преимущество правого поля зрения, т. е. левого полушария [31, 32]. Авторы этих работ также считают, что* в правом полушарии осуществляется преимущественно зрительно-пространственный анализ, физических параметров вербальных стимулов, а в левом полушарии-—специфических свойств, связанных непосредственно с речевой функцией. Если это так, то функция зрительного восприятия, словесных сигналов полноценно может осуществляться при непременном взаимодействии двух полушарий. И действительно, при поражениях правого полушария существенно нарушается опознание неполно или нечетко обрисованных букв [33], речь становится маловыразительной, «менее личностной», нарушается дифференцирование фонем [34, 35], расстраивается лроцесс непроизвольного запоминания слов [36].
Концепция о взаимодополняющем «сотрудничестве» двух полушарий и преимущественно отдельного полушария в осуществлении лишь определенной стадии той или иной нервно-психической деятельности, а не всей функции в целом подтверждается также путем сравнения эффективности выполнения задачи при адресовании словесных стимулов одновременно в оба полушария и в одно полушарие, левое или правое. При подаче информации в оба полушария задача выполняется значительно эффективнее [37].
О значении правого полушария в восприятии вербальной информации говорят также результаты, полученные О. А. Генкиной при регистрации вызванных потенциалов у лиц с патологически низким интеллектуальным уровнем. У этого рода исследуемых не отмечается вышеупомянутого «опережения» в правом полушарии коркового вызванного ответа. Мы думаем, что это является одним из нервных механизмов нарушений у олигофренов процессов переработки речевой информации.
Таким образом, можно думать, что в едином и сложном нервно-психическом процессе речевого восприятия его начальная стадия в норме осуществляется при непременном участии правого полушария.
ЛИТЕРАТУРА
Bryden M. P. Neuropsychologia, 1965, v. 3, p. 1.
Bryden M. P. Cortex, 1973, v. 9, No. 4, p. 419.
Fontenot D. J. J. Compar. and Physiol. Psychol., 1973, v. 85, No. 3, p. 564.
Kimura D. Sclent. Amer., 1973, v. 228, No. 3, p. 70.
White M. J. Psychol. Bull, 1969, v. 72, No. 6, p. 387.
White M. J. J. Exptl Psychol., 1970, v. 83, No. 1, p. 174.
White M. J. Canad. J. Psvchol., 1971, v. 25, p. 207.
White M. J. Quart. J. Exptl Psychol., 1973, v. 25, p. 41.
Sperry R. W. In: Brain and conscious experience. Berlin — Heidelberg — New York,1966, 298 p.
Gazzaniga M. S., Hillyard S. A. Neuropsychologia, 1971. v. 9, No. 3, p. 273.
Gazzaniga M. S., Boden J. E., Sperry R. W. Brain, 1965, v. 88, p. 221.
Глезер В. Д., Дудкин К- Н., Куперман А. М, Леушина Л. И., Невская А. А., Подви-
гин Н. Ф., Праздникова Н. В. В кн.: Зрительное опознание и его нейрофизиологиче
ские механизмы. Под ред. В. Д. Глезера. Л., «Наука», 1975, с. 269.
Lindsley D. В., Emmons W. Н. Science, 1958, v. 127, No. 330, p. 1061.
Battersley W. S., Wagman I. H. Amer. J. Physiol., 1962, v. 203, p. 359.
Raab D. H. Psychol. Bull., 1963, v. 60, p. 118.
Matsumiya Y., Tagliasco V., Zombroso С. Т., Goodglass H. Science, 1972, v. 175,
No. 4023, p. 790.
Morrell L. K., Salamy J. G. Science, 1971, v. 174, No. 4005, p. 164.
Wood Ch. C, Goff W. R., Day R. S. Science, 1971, v. 173, No. 4003, p. 1248.
Rattiff S. S., Greenberg H. J. J. Auditory Res., 1972, v. 12, No 1, p. 14
Shelburne S. A. EEG and Clin. Neurophysiol., 1972, v. 32, No. 1, p. 17.
Buchsbaum M., Fedio P. Physiol. and Behaviour, 1970, v. 5, p. 207.
Gott R. S., Boyarsku L. L. J. Neurobiol., 1972, v. 3, No. 1, p. 66.
Buchsbaum M., Fedio P. EEG and Clin. Neurophysiol., 1969, v. 26, No. 3, p. 266.
Газанига М. В кн.: Восприятие. Механизмы и модели. М., «Мир», 1974, с. 45.
Mishkin M. In: Brain and Human Behaviour (A. G. Karczmar, J. C. Eccles, eds.). Ber
lin — Heidelberg — New York, 1972, p. 187.
Ritter W., Simson R., Vaughan H. G. EEG and Clin. Neurophysiol., 1972, v. 33, No. 6,
p. 547.
:27. Zangwill O. Z. Cerebral dominance and its relations to psychological function. Edinburgh, Oliver and Boyd, 1960.
Yeni-Komshian G. H., Isenberg D., Goldberg H. Neuropsychologia, 1975, v. 13, No. 1,
p. 83.
Rhodes L. E., Dustman R. S., Beck E. C. EEG and Clin. Neurophysiol., 1969, v. 27, p. 364.
30. Bigum H. В., Dustman R. S., Beck E. C. EEG and din! Neurophysiol., 1970, v. 28,
No. 6, p. 576.
Cohen G. In: Attention and peilormance. (P. M. A. Rabbit, S. Darnic, eds). London,
Acad. Press, 1975, p. 20.
Geffen G., Bradshaw J. L., Nettleton N. C. J. Exptl Psychol., 1972, v. 95, No. 1, p. 25.
Warringtoh E. K-, James M. Neuropsychologia, 1967, v. 5, No. 3, p. 253.
Лебединский М. С. В кн.: Проблемы современной психиатрии. М., Изд-во АМН
СССР, 1948, с. 85.
Лебединский М. С. Ж- невропатол. и психиатрии, 1964, т. 64, вып. 1, с. 36.Лурия А. Р., Симерницкая Э. Г. Физиол. человека, 1975, т. 1, № 3, с. 411.
-37. Dimond S., Beaumont G. Nature, 1971, с. 232, No. 5308, p. 270.








Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!

